
Napriek desaťročiam štúdií zostávajú molekulárne mechanizmy a selektivita biomolekulárnych zložiek jedu včiel medonosných (Apis mellifera) ako protirakovinových látok do značnej miery neznáme. Tu demonštrujeme, že včelí jed a jeho hlavná zložka melitín silne indukujú bunkovú smrť, najmä v agresívnych trojnásobne negatívnych a HER2 obohatených podtypoch rakoviny prsníka. Včelí jed a melitín potláčajú aktiváciu EGFR a HER2 tým, že interferujú s fosforyláciou týchto receptorov v plazmatickej membráne buniek karcinómu prsníka. Mutačné štúdie odhaľujú, že pozitívne nabitá C-terminálna sekvencia melittínu sprostredkúva interakciu plazmatickej membrány a protirakovinovú aktivitu. Vytvorenie motívu RGD ďalej zvyšuje zacielenie melitínu na malígne bunky s minimálnou toxicitou pre normálne bunky. Nakoniec podávanie melittínu zvyšuje účinok docetaxelu pri potlačovaní rastu nádoru prsníka v modeli aloštepu. Naša práca odhaľuje molekulárny mechanizmus podporujúci protirakovinovú selektivitu melittínu a načrtáva liečebné stratégie zamerané na agresívne rakoviny prsníka.
Úvod
Európska včela medonosná (Apis mellifera) je už tisícročia zdrojom množstva produktov používaných v medicíne, ako je med, propolis a jed /1/. Molekulárne determinanty protirakovinovej aktivity včelieho jedu však zostávajú nedostatočne pochopené, najmä v prípade rakoviny prsníka, najbežnejšej rakoviny u žien na celom svete /2/. Pochopenie molekulárneho základu a špecifickosti včelieho jedu proti rakovinovým bunkám je kľúčom k vývoju a optimalizácii nových účinných terapeutík z prírodného produktu, ktorý je široko dostupný a nákladovo efektívny na výrobu v mnohých komunitách po celom svete.
Aktívnou zložkou včelieho jedu je melittín, ktorý obsahuje polovicu včelieho jedu v sušine /3,4/. Melittín je pozitívne nabitý amfipatický 26-aminokyselinový peptid5, ktorý sa spája s fosfolipidmi membránovej dvojvrstvy a spôsobuje bunkovú smrť vytvorením transmembránových toroidných pórov s priemerom ~4,4 nm, ktoré môžu umožniť internalizáciu ďalších malých molekúl s cytotoxickými aktivitami /4,6 ,7/.
Včelí jed aj melitín preukázali protinádorové účinky u melanómu8, nemalobunkového karcinómu pľúc /9/, glioblastómu /10/, leukémie /11/, vaječníkov /12/, krčka maternice /13/ a rakoviny pankreasu /14/, s vyššou cytotoxickou účinnosťou v rakovinových bunkách v porovnaní s netransformovanými bunkami /8,11,14,14,14,14,/ Nanočastice melittínu sa používajú na potlačenie pečeňových metastáz prostredníctvom imunomodulácie pečeňových sínusoidných endotelových buniek /16/. Medzi včelím jedom a inými terapeutickými spôsobmi, vrátane cisplatiny pri cervikálnych a laryngeálnych malignitách /17/, a docetaxelu v bunkách rakoviny pľúc, boli hlásené aditívne a synergické protirakovinové účinky. Podobné interakcie boli preukázané medzi melitínom a plazmou ošetreným fosfátom pufrovaným fyziologickým roztokom v bunkách rakoviny prsníka a melanómu MCF719. Včelí jed a melitín tiež indukovali apoptózu v bunkách MCF720 a znížili životaschopnosť a migráciu buniek v bunkách rakoviny prsníka MDA-MB-231 /21,22/. Včelí jed znížil metastázy rakoviny prsníka do pľúc /23/, inhiboval rast nádoru a predĺžil prežitie u myší so spontánnymi nádormi prsníka /24/. Väčšina antineoplastickej aktivity včelieho jedu sa pripisuje melittínu /25/ prostredníctvom inhibície osi PI3K/Akt/mTOR pri rakovine prsníka21, MAPK pri melanóme /26/, JAK2/STAT3 pri rakovine vaječníkov /12/ a signálnych dráh NFκB v bunkách karcinómu pľúc /18/. Na rozdiel od včelieho jedu neobsahuje jed čmeliaka (Bombus terrestris) melittín /27/, ale obsahuje sekrečnú fosfolipázu A2, ktorá indukovala apoptózu inhibíciou fosforylácie Akt v bunkách ľudskej chronickej myeloidnej leukémie /28/.
Podľa našich najlepších vedomostí účinky rôznych včelích jedov a melittínu naprieč podtypmi rakoviny prsníka v porovnaní s netransformovanými bunkami neboli skúmané. Triple-negatívne rakoviny prsníka (TNBC, ktorým chýba expresia estrogénových a progesterónových receptorov, ako aj receptora ľudského epidermálneho rastového faktora 2, HER229) sú agresívne a spájajú sa s najhoršími výsledkami /30,31,32,33/. Približne 50 % TNBC nadmerne exprimuje receptor epidermálneho rastového faktora (EGFR) /34/ a nádory obohatené o HER2 nadmerne exprimujú HER2, ďalšiu receptorovú tyrozínkinázu (RTK), ktorá prepožičiava onkogénnu signalizáciu často závislú od dráhy PI3K/Akt downstream /34/. Blokovanie signalizácie EGFR v TNBC pomocou štandardných terapií preukázalo obmedzenú klinickú účinnosť v skorých fázach klinických štúdií v dôsledku nedostatočnej závislosti na dráhe EGFR a dôležitosti kolaterálnych dráh /35/. Hoci terapie zamerané na HER2 dramaticky zlepšili medián prežitia v metastatickom prostredí, rezistencia je pre tento podtyp takmer nevyhnutná aj z dlhodobého hľadiska /33,36/. Je zrejmé, že objavenie účinnejších a selektívnejších terapeutických stratégií pre tieto druhy rakoviny je prioritnou oblasťou v klinickej onkológii.
Tu uvádzame, že včelí jed a melitín indukujú silnú a vysoko selektívnu bunkovú smrť v TNBC a HER2 obohatenom karcinóme prsníka so zanedbateľnými účinkami v normálnych bunkách, tým, že interferujú s RTK interakciami závislými od rastového faktora kritickými pre fosforyláciu receptora a aktiváciu PI3K/Akt signalizácia. Okrem rakoviny prsníka uvádzame aj cielené modifikácie melittínu na potenciálne použitie v kombinácii s chemoterapiou na liečbu iných agresívnych rakovín poháňaných nadmernou expresiou receptorov rastových faktorov.
Výsledky
Včelí jed a melittín znižujú životaschopnosť rakoviny prsníka
Na posúdenie protirakovinovej účinnosti a selektivity sa jed z oboch európskych včiel zozbieraných v Perthe v Austrálii a melitínový peptid hodnotili v testoch dávka-odpoveď v paneli bunkových línií reprezentatívnych pre vnútorné podtypy rakoviny prsníka av netransformovaných bunkách (obr. 1a). Včelí jed vykazoval vysokú protirakovinovú selektivitu s výrazne vyššou účinnosťou v TNBC (napr. SUM159 a SUM149) a v bunkových líniách rakoviny prsníka obohatených HER2 (napr. MDA-MB-453 a SKBR3), po ktorých nasledovali luminálne bunky rakoviny prsníka ( vrátane MCF7 a T-47D), s najnižším vplyvom na normálne bunky (primárne dermálne fibroblastové bunky HDFa a prsné netransformované bunky MCF 10A a MCF-12A) (obr. 1b, vľavo; tabuľka 1; GLM, Wald Chi-Square = 342, p < 0,001, n = 33, df = 1). V porovnaní s normálnou bunkovou líniou HDFa (22,17 ng/) sa pozorovalo významné zníženie polovičnej maximálnej inhibičnej koncentrácie (IC50) pre rakovinové bunkové línie TNBC SUM159 (5,58 ng/μL) aj HER2 obohatené SKBR3 (5,77 ng/μL). 1c, vľavo, jednosmerná ANOVA, p < 0,01).

Tabuľka 1 Polovičné maximálne inhibičné koncentrácie (IC50) včelieho jedu a melitínu.

Podobne bol melitín významne účinnejší proti HER2-obohatenej rakovine prsníka a TNBC v porovnaní s normálnymi bunkami (obr. 1b, c, vpravo; tabuľka 1; GLM, Wald Chi-Square = 12,9, p < 0,001, n = 33, df = 1), s hodnotami IC50 od 0,94 do 1,49 μM v ľudských bunkách rakoviny prsníka obohatených o TNBC a HER2 a 1,03 až 2,62 μM v netransformovaných bunkách. Testy bunkovej životaschopnosti včelieho jedu a melitínu v myšacej rakovine prsníka a normálnych bunkových líniách potvrdili zvýšenú selektivitu pre agresívne myšacie nádorové bunkové línie, ako je p53-mutant claudín-low T11 a BRCA-mutant B.1537,38 (doplnkový obrázok 1).
Jed včiel z rôznych populácií včiel v Írsku a Anglicku znížil životaschopnosť buniek SUM159 a SKBR3 podstatne viac ako netransformovaných buniek HDFa (obr. 1d, jednosmerná ANOVA, p < 0,001). Testovali sme aj jed čmeliaka Bombus terrestris z Anglicka. Vzorky od robotníc aj kráľovien vyvolali minimálnu bunkovú smrť v bunkách rakoviny prsníka v porovnaní s jedom včiel aj pri vysokých koncentráciách jedu (obr. 1e).
Vyvinuli sme myšaciu monoklonálnu protilátku rozpoznávajúcu melitín na posúdenie relatívneho množstva melitínu vo všetkých vzorkách jedu včiel a čmeliakov pomocou ELISA. V súlade s vyššie uvedenými štúdiami aktivity sa relatívna abundancia melitínu významne nelíšila vo všetkých vzorkách včelieho jedu z rôznych miest (obojsmerná ANOVA, p > 0,999). Avšak koncentrácie melittínu boli významne vyššie vo vzorkách včiel v porovnaní s jedom čmeliaka a izotypovou IgG kontrolou (obr. 1f, dvojcestná ANOVA, p < 0,001).
Protirakovinové účinky melitínu boli potvrdené blokovacími experimentmi in vitro, v ktorých sme využili anti-melitínovú protilátku na záchranu životaschopnosti buniek v bunkách HDFa a SUM159. Bunky boli ošetrené včelím jedom alebo melitínom v kombinácii so zvyšujúcimi sa koncentráciami protilátky proti melitínu. Životaschopnosť buniek bola signifikantne vyššia, keď bol melitín blokovaný protilátkou proti melitínu pre bunky HDFa a SUM159 vystavené včelímu jedu alebo peptidu melitínu (obr. 1g, t testy, p < 0,0001). Tieto údaje naznačujú, že melitín prítomný v jede včiel je najvýznamnejšou bioaktívnou protirakovinovou zlúčeninou spomedzi všetkých študovaných jedov. Na všetky ďalšie experimenty sa použil včelí jed zozbieraný v austrálskom Perthe.
Včelí jed a melittín spôsobujú smrť buniek rakoviny prsníka
Na preskúmanie mechanizmu a kinetiky bunkovej smrti boli bunky TNBC ošetrené IC50 buď včelím jedom alebo melittínom počas 18 a 24 hodín a spracované testom štiepenej kaspázy-3 na kvantifikáciu apoptotickej bunkovej smrti. Imunoblotting potvrdil indukciu štiepenej kaspázy-3 v bunkách SUM159, pričom samotný melitín indukoval vyššiu úroveň apoptózy ako včelí jed 18 aj 24 hodín po ošetrení (obr. 2a, kvantifikácia na doplnkovom obrázku 2).
Obr. 2: Včelí jed a melittín indukujú apoptózu a narušenie membrány.

Na kvantifikáciu apoptotických, nekrotických alebo mŕtvych bunkových populácií po liečbe sme vykonali test detekcie apoptózy Annexin V-FITC. Bunky SUM159 boli vystavené pôsobeniu vehikula, včelieho jedu alebo melitínu s použitím koncentrácií IC50 a spracované prietokovou cytometriou po 60-minútovom ošetrení (obr. 2b). Zistili sme signifikantne viac neskorých apoptotických/nekrotických buniek pre vzorky ošetrené melitínom (23,6 ± 5,7 %) v porovnaní s včelím jedom (8,3 ± 1,9 %) a kontrolou s vehikulom (4,8 ± 0,4 %, obojsmerná ANOVA, p 01 , priemer 0. ± SEM). Neexistovali však žiadne významné rozdiely v hladinách skorých apoptotických alebo nekrotických buniek vo všetkých podmienkach (dvojcestná ANOVA, p > 0,05, priemer ± SEM). Aby sa charakterizovala kinetika bunkovej smrti v kratších časoch, merala sa životaschopnosť buniek pre bunky HDFa, SKBR3 a SUM159 ošetrené až 1 hodinu s koncentráciami IC50 včelieho jedu alebo melitínu (obr. 2c). Včelí jed rapídne znížil životaschopnosť buniek bez významného rozdielu medzi normálnymi a rakovinovými bunkovými líniami v priebehu hodiny (obojsmerná ANOVA, p = 0,97). Na rozdiel od toho melitín významne znížil životaschopnosť oboch bunkových línií rakoviny prsníka v porovnaní s normálnymi bunkami od 10 min ďalej a SUM159 významne viac ako SKBR3 od 30 min ďalej (dvojcestná ANOVA, p < 0,0001).
Konfokálna mikroskopia živých buniek (obr. 2d) a skenovacia elektrónová mikroskopia (obr. 2e) v bunkách SKBR3 a SUM159 ukázali rýchle narušenie a zmršťovanie plazmatickej membrány pomocou včelieho jedu a melitínu v porovnaní s liečbou vehikulom počas 10 až 60 minút.
RGD zvyšuje zacielenie melittínu na rakovinu prsníka
C-koniec melitínu tvorí kladne nabitú a-helix, o ktorej sa predpokladá, že sprostredkuje väzbu na negatívne nabitú plazmatickú membránu, čo vyvolá následnú tvorbu pórov a lýzu buniek39, 40, 41. Predchádzajúce štúdie ukázali, že skrátenie tohto pozitívne nabitého C-konca významne znižuje väzbu melitínu na fosfolipidové dvojvrstvy v porovnaní s melitínom divokého typu39,42. Na posúdenie funkčnej úlohy pozitívnej (K21RKR24) sekvencie na C-konci melitínu sme navrhli negatívne nabitý melitínový peptid (D21EDE24-melitín). Predpokladalo sa, že tieto negatívne zvyšky narušia väzbu melitínu s plazmatickou membránou. Zistili sme, že DEDE-melitín nevyvoláva žiadne merateľné známky protirakovinovej aktivity v žiadnej z testovaných bunkových línií (obr. 3a, b). Dôležité je, že protirakovinová aktivita DEDE-melittínu bola zachránená pozitívne nabitou sekvenciou (K21KKRKV26) prítomnou vo veľkom T antigéne opičieho vírusu 40 (SV40) (peptid SV40-melittín), ktorý má schopnosť prenikať do buniek43 (obr. 3b). Podobne vrúbľovanie väčšej pozitívne nabitej sekvencie TAT (transaktivátor transkripcie, odvodený z HIV-1)43 na C-koniec melitínu tiež obnovilo aktivitu DEDE-melitínu (peptid TAT-melittín; doplnkový obrázok 3). Avšak sila melittínu a SV40-melittínu bola väčšia ako TAT-melittín, čo mohlo byť spôsobené väčšou veľkosťou TAT. Tieto údaje demonštrujú, že zvyšky potrebné pre aktivitu melitínu zahŕňajú zvyšky nachádzajúce sa v C-terminálnom a-helixe, ktorý obsahuje niekoľko kľúčových kladne nabitých zvyškov potrebných na interakciu s plazmatickou membránou.
Obr. 3: Technika melitínu s motívom RGD zvyšuje selektivitu rakoviny prsníka.
Na zvýšenie selektivity rakovinových buniek sme vytvorili bifunkčný melitínový peptid skonštruovaním N-koncového alfa-helikálneho peptidového motívu RGD (RGD1-melittín, odvodený z TGF-p3, sekvencia HGRGDLGRLKK), ktorý interaguje s integrínmi avp6 a avp3 nadmerne exprimovanými pri rakovine prsníka bunkové membrány a vaskulatúra spojená s nádorom44,45,46. Keď sú RGD motívy upravené pomocou bioaktívnych peptidov, zvyšujú zacielenie na bunky rakoviny prsníka47. Hodnota IC50 RGD1-melitínu sa významne nelíšila v porovnaní s rodičovským melittínom v bunkách T11, čo naznačuje, že účinnosť nebola ovplyvnená motívom RGD (obr. 3b, t test, p = 0,652). Ak vezmeme do úvahy pomery IC50 HDFa/SUM159 pre RGD1-melitín (2,73 ± 0,14) v porovnaní s melitínom (1,76 ± 0,04), motív RGD významne zvýšil terapeutické okno medzi normálnymi a TNBC bunkovými líniami, čo potvrdzuje zvýšenú selektivitu rakovinových buniek pomocou RGD (obr. 3c, t test, p < 0,01, priemer ± SEM). Indukcia apoptózy v SUM159 TNBC bunkách ošetrených melitínom, DEDE-melitínom a RGD1-melitínom počas 24 hodín potvrdila protirakovinovú aktivitu melitínu aj RGD1-melitínu, ale nie DEDE-melitínu (obr. 3d).
V súlade s protirakovinovou aktivitou melitínu a RGD1-melitínu sme zistili, že interakcia medzi anti-melitínovou protilátkou a melitínom sa významne nelíšila od interakcie s RGD1-melitínom (obr. 3e, dvojcestná ANOVA, p > 0,999), ale bola významne odlišná od DEDE-melittínu a SV40-melitínu (dvojcestná ANOVA, p < 0,05), pričom absorbancia SV40-melitínu sa významne nelíšila od IgG kontroly (dvojcestná ANOVA, p > 0,1). Tieto údaje naznačujú, že naša monoklonálna anti-melitínová protilátka rozpoznáva konformačný epitop, ktorý nie je narušený skonštruovaním N-koncového zameriavacieho peptidu.
Modelovacie štúdie ukázali, že konformácia melitínovej časti upravených peptidov nebola narušená ani C-terminálnymi mutáciami, ani N-terminálnou adíciou RGD motívu (obr. 3f). Každý peptid si zachoval charakteristickú ohnutú štruktúru alfa-helixu, ktorá potenciálne uľahčuje tvorbu pórov4, čo naznačuje, že rozdiely v protirakovinovej aktivite medzi mutantmi sú spôsobené elektrostatickými interakciami s membránou a nie veľkými zmenami v štruktúre peptidu.
Ďalej sme použili anti-melitínovú protilátku na detekciu subcelulárnej lokalizácie aktívnych peptidov imunofluorescenciou v TNBC SUM159 bunkách ošetrených 30 min vehikulom, včelím jedom, melitínom, RGD1-melitínom alebo DEDE-melittínom v koncentráciách IC50 (obr. 3g ). Nezávisle od toho, či boli bunky vystavené včelímu jedu, melitínu alebo RGD1-melitínu, melitín bol lokalizovaný prevažne na plazmatickej membráne buniek nadmerne exprimujúcich EGFR, so stupňom intracelulárneho zafarbenia v včelím jedu a bunkách ošetrených melitínom, potenciálne v dôsledku narušenia membrány a tvorba endozómov, ako sa uvádza inde25,48. Okrem toho sa vzor farbenia RGD1-melitínu javil ako zreteľne zameraný na plazmatickú membránu samotnú, čo by bolo v súlade so zvýšenou selektivitou cieľového peptidu pre časti povrchu nádorových buniek. Pozorovali sme nedostatok reaktivity melitínovej protilátky v bunkách ošetrených DEDE-melitínom. V súhrne tieto výsledky ukazujú, že zatiaľ čo motív RGD zvyšuje zacielenie melitínu na bunkové membrány rakoviny prsníka, pozitívny motív na C-konci sa zdá byť nevyhnutný pre protirakovinovú aktivitu.
Včelí jed a melitín potláčajú fosforyláciu RTK
Následne sme skúmali, či včelí jed aj melitín narušujú signálne dráhy spojené s RTK blokovaním ligandovo závislej aktivácie EGFR a HER2 v bunkách karcinómu prsníka. Aby sme to zhodnotili, vykonali sme imunoblotovaciu analýzu na extraktoch SKBR3 (HER2+ a EGFR+) a SUM159 (EGFR+) buniek vystavených EGF a ošetrených IC50 včelieho jedu alebo melitínu od 2,5 do 20 minút (obr. 4a). Včelí jed aj melitín znižovali fosforyláciu RTK a modulovali súvisiace signálne dráhy PI3K-/Akt a MAPK spôsobom závislým od času.
Obr. 4: Včelí jed a melitín potláčajú fosforyláciu EGFR a HER2.
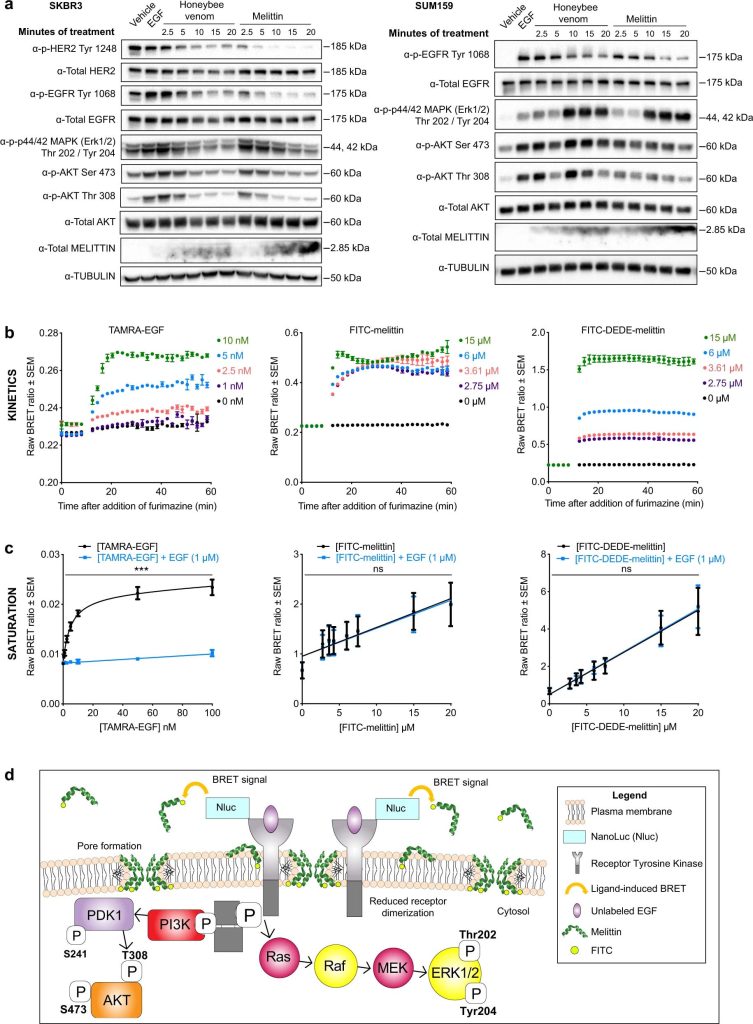
Liečba včelím jedom a melitínom v bunkách SKBR3 výrazne znížila p-HER2 (Tyr1248), p-EGFR (Tyr1068), p-p44/42 MAPK (Thr202/Tyr204), p-Akt (Ser473 a Thr308), p-SAPK/JNK (Thr183/Tyr185) a p-p38 MAPK (Thr180/Tyr182) od 5 min ďalej (obr. 4a, vľavo; doplnkový obrázok 4), s miernym poklesom celkového proteínu HER2, EGFR a Akt iba po 10 min. liečby včelím jedom, čo by mohlo súvisieť s degradáciou receptorov sprostredkovanou endozómami25. V SUM159 bol p-EGFR (Tyr1068) silne downregulovaný včelím jedom a melitínom od 10 do 20 minút. Liečba SUM159 melitínom tiež potlačila p-Akt (Ser473 a Thr308) vo všetkých časových bodoch, ale zvýšila reguláciu p-p44/42 MAPK (Thr202/Tyr204), p-SAPK/JNK (Thr183/Tyr185) a p-p38 MAPK ( Thr180/Tyr182) od 10 do 20 min, zatiaľ čo jed včiel zvýšil reguláciu p-p44/42 MAPK (Thr202/Tyr204) a p-Akt (Ser473 a Thr308) z 10 na 20 min; Obr. 4a, vpravo ). Dráhy MAPK a Akt mohli byť v bunkách SUM159 upregulované v dôsledku uvoľnenia negatívnej regulačnej spätnoväzbovej slučky, ktorá spúšťa signalizáciu ERK na ochranu buniek pred apoptotickou bunkovou smrťou8,49. Anti-melitínová protilátka indikovala zvyšujúce sa množstvo melitínu prítomného v lyzátoch oboch bunkových línií v priebehu času, so silnejším signálom pre ošetrenie melitínom v porovnaní s včelím jedom v oboch bunkových líniách.
Aby sme charakterizovali účinky na signálne dráhy v inom modeli TNBC, uskutočnili sme imunoblotting na bunkách MDA-MB-231, v ktorých liečba EGF fosforylovala EGFR a vyvolala expresiu EGFR (doplnkový obrázok 4). Melittín znižoval fosforyláciu EGFR a MAPK, čím nadol reguloval hlavné dráhy onkogénnej proliferácie. Na rozdiel od buniek SUM159 stimulácia EGFR pomocou EGF nekorelovala so zvýšením fosforylácie v p-Akt, potenciálne v dôsledku odpojenia medzi signalizáciou EGFR a dráhami Akt. Iné receptory rastových faktorov, ako je VEGFR1, môžu sprostredkovať aktiváciu týchto dráh50,51. Zatiaľ čo melitín predtým inhiboval signalizáciu JAK2 / STAT3 pri rakovine vaječníkov12, nepozorovali sa žiadne modulačné účinky na inhibítory dráhy JAK / STAT v bunkách SUM159 po 60-minútovom ošetrení včelím jedom alebo melitínom (doplnkový obrázok 5).
Vzhľadom na to, že bunky karcinómu prsníka obohatené o TNBC a HER2 sú vysoko závislé od aktivácie EGFR a HER2, uskutočnili sme experimenty s bioluminiscenčným rezonančným prenosom energie (BRET), aby sme určili, či melitín interferuje s väzbou EGF na EGFR, čo vedie k pozorovanému potlačeniu rastu. fosforylácia receptora faktora. Reportér NanoLuc sa použil ako bioluminiscenčná donorová molekula a geneticky sa spojil s EGFR52,53. Kinetické a saturačné BRET experimenty sa použili na monitorovanie blízkosti NanoLuc-EGFR s fluorescenčne označenými akceptorovými molekulami TAMRA-EGF (pozitívna kontrola), FITC-melitín a FITC-DEDE-melitín (negatívna kontrola) v bunkách HEK293FT transfekovaných NanoLuc- EGFR. K prenosu energie z bioluminiscenčného donoru na fluorescenčný akceptor dochádza na vzdialenosti menšie ako 10 nm a svedčí o interakciách medzi sledovanými označenými molekulami54. Signál BRET sa určuje monitorovaním pomeru emisie svetla z akceptora k emisii z darcu.
Vybral sa rozsah koncentrácií každého peptidu, vrátane IC50 FITC-melittínu, so zodpovedajúcimi molárnymi koncentráciami FITC-DEDE-melitínu. Zistili sme, že signál BRET sa zvýšil v závislosti od dávky pre TAMRA-EGF a FITC-DEDE-melitín a v menšej miere pre FITC-melitín (obr. 4b). FITC-DEDE-melitín vykazoval oveľa vyššie pomery BRET ako FITC-melitín pri rovnakých koncentráciách, ako aj veľmi rýchlo dosahoval maximálne pomery BRET pri každej dávke. Nešpecifický peptid navrhnutý proti transkripčnému faktoru Engrailed 1 (EN1)55 (FITC-EN1-mutant) vykazoval podobné pomery BRET a kinetiku ako FITC-DEDE-melitín (doplnkový obrázok 6), čo naznačuje, že na zistenie špecificity boli potrebné ďalšie experimenty. väzbové interakcie s EGFR.
Aby sme určili špecificitu väzby melittínu na EGFR na väzbovom mieste EGF, uskutočnili sme saturačné BRET testy na vyhodnotenie kompetície EGF s každým z peptidov viažucich sa na NanoLuc-EGFR (obr. 4c). Zatiaľ čo väzba TAMRA-EGF na NanoLuc-EGFR bola saturovateľná a významne znížená v prítomnosti 1 uM EGF (dvojcestná ANOVA, p < 0,0001), signály BRET melitínu FITC a melitínu FITC-DEDE neboli saturovateľné a sa významne nelíšia s alebo bez 1 uM EGF (dvojcestná ANOVA, p > 0,999), čo naznačuje, že ani melitín, ani DEDE-melitín sa neviažu na EGF-väzbové miesto.
Naše údaje podporujú názor, že melitín sa začleňuje do plazmatickej membrány rakovinových buniek prostredníctvom nabitej sekvencie prítomnej na C-konci, čo vyvoláva remodeláciu a rozrušenie plazmatickej membrány. Údaje BRET naznačujú, že melitín môže byť umiestnený vo vzdialenosti do 10 nm od RTK bez toho, aby interferoval s väzbovým miestom endogénneho rastového faktora (obr. 4d).
Melittín senzibilizuje TNBC na liečbu docetaxelom in vivo
Ďalej sme testovali potenciálne synergie medzi melitínom a chemoterapeutickými látkami na zvýšenie smrti buniek rakoviny prsníka. Myšia bunková línia p53-TNBC T11 bola ošetrená docetaxelom v kombinácii buď s včelím jedom alebo melitínom a na stanovenie kombinačného indexu (CI) medzi ošetreniami boli uskutočnené testy bunkovej životaschopnosti56 (obr. 5a). Pozorovali sme CI < 1 pre všetky testované koncentrácie, čo naznačuje silné synergické interakcie (obr. 5b). Synergizmy sa pozorovali aj s cisplatinou, látkou používanou na liečbu TNBC na klinike (doplnkový obrázok 7). Model xenoštepu T11 sa použil na experimenty in vivo, pretože preukázal najpriaznivejšiu liekovú interakciu in vitro medzi melitínom a docetaxelom naprieč viacerými testovanými bunkovými líniami (doplnkový obrázok 8) a má intaktný imunitný systém umožňujúci imunitnú odpoveď na melitín posúdiť.
Obr. 5: Melittín senzibilizuje vysoko agresívne nádory TNBC na liečbu docetaxelom in vivo.
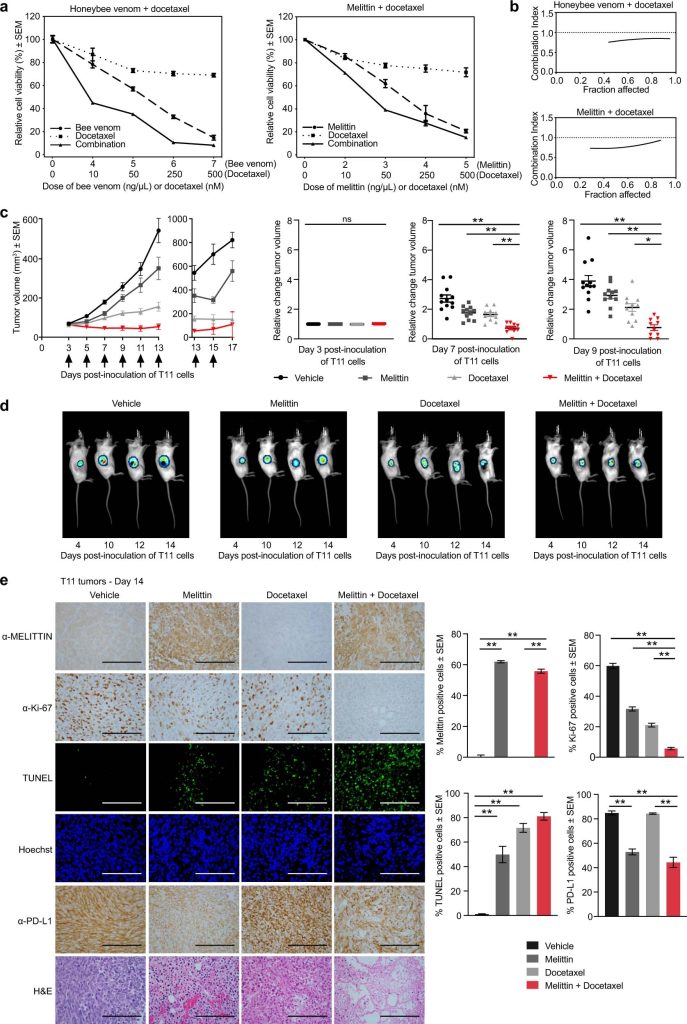
Aby sme preskúmali účinnosť kombinácie melitínu a docetaxelu pri znižovaní rastu TNBC, uskutočnili sme in vivo experimenty transplantáciou buniek T11 do myší BALB/c. Tento model aloštepu rekapituluje vysoko agresívne ochorenie TNBC s nízkym obsahom klaudínu u myší s intaktným imunitným systémom38, 57, 58. Tri dni po vytvorení nádorov T11 (~50 mm3) boli myši náhodne rozdelené do štyroch skupín (n = 12 myší/skupina) a intratumorálne liečené vehikulom, melitínom (5 mg/kg), docetaxelom (7 mg/kg), alebo kombinácia melittínu (5 mg/kg) a docetaxelu (7 mg/kg). Myši boli liečené každé 2 dni od 3. dňa, celkovo 7 liečení. Zistili sme, že pri kombinovanej liečbe bola kontrola nádoru lepšia v porovnaní s liečbou samotnou alebo vehikulom, najmä v dňoch 7 a 9 po inokulácii buniek T11, pričom táto kombinácia dosiahla významné zníženie objemu nádoru (obr. 5c, jednosmerný ANOVA, p < 0,001). To naznačuje, že nádory rezistentné na docetaxel by sa mohli stať citlivými pridaním melitínu. Tieto štúdie sme overili bioluminiscenčným zobrazovaním (BLI), aby sme neinvazívne sledovali zmeny v raste nádoru in vivo v bunkách T11 označených konštruktom obsahujúcim luciferázu (obr. 5d). Tu sme opäť zistili zlepšenú kontrolu nádoru pri kombinovanej liečbe docetaxelom a melittínom na 10., 12. a 14. deň v porovnaní so všetkými ostatnými skupinami.
Terapeutické účinky melitínu a docetaxelu boli overené v nádorových tkanivách 14. deň po inokulácii buniek T11 imunohistochémiou a imunofluorescenciou (obr. 5e). Protilátka proti melitínu potvrdila intratumorálnu lokalizáciu buniek pozitívnych na melitín v skupine s melitínom (61,9 ± 0,7 %) aj v skupine s kombinovanou liečbou (55,8 ± 1,3 %), ale nie v kontrolnej skupine s vehikulom (jednosmerná ANOVA, p. < 0,01, priemer ± SEM). Významné zníženie proliferácie nádorových buniek (hodnotené expresiou Ki-67) sa zistilo v nádoroch liečených kombináciou melitínu a docetaxelu (5,7 ± 0,8 %) v porovnaní s vehikulom (59,8 ± 1,7 %) v porovnaní s ktorýmkoľvek melitínom ( 31,7 ± 1,3 %) alebo samotný docetaxel (21,0 ± 1,3 %, jednosmerná ANOVA, p < 0,01, priemer ± SEM). Farbenie TUNEL potvrdilo významne vyššiu fragmentáciu DNA a indukciu apoptózy v skupine s kombináciou (81,0 ± 3,1 %) v porovnaní s vehikulom (1,0 ± 0,4 %, jednosmerná ANOVA, p < 0,01, priemer ± SEM).
Ligand-1 programovanej smrti proteínu imunitného kontrolného bodu (PD-L1) znižuje funkčnosť aktivovaných T buniek. V dôsledku toho blokády imunitných kontrolných bodov v kombinácii s chemoterapiou zabraňujú rozpoznaniu PD-L1 T-buniek, čím zabraňujú tejto adaptívnej imunitnej rezistencii v TNBC, a tým zvyšujú terapeutickú účinnosť oproti samotnej chemoterapii59. Na rozdiel od samotného docetaxelu (84,3 ± 0,6 %), ktorý neovplyvnil hladiny PD-L1 v nádoroch, zistili sme, že melittín významne znížil expresiu PD-L1 v nádoroch, keď sa použil samostatne (52,9 ± 2,4 %) alebo v kombinácii liečba (44,3 ± 4,2 %) v porovnaní s vehikulom (84,9 ± 1,6 %, jednosmerná ANOVA, p < 0,01, priemer ± SEM). Stručne povedané, tieto štúdie podporujú názor, že melitín senzibilizuje bunky T11 na liečbu docetaxelom a že melitín by mohol pomôcť zmierniť expresiu proteínov kontrolných bodov imunity, čím by sa následne zlepšili protinádorové imunitné reakcie.
Ďalej sme vykonali imunohistochémiu v liečených nádoroch T11 na detekciu p-HER2 (Tyr1248) a p-EGFR (Tyr1068) (doplnkový obrázok 9). Expresia EGFR bola mierne, ale významne znížená kombináciou melittínu a docetaxelu (75,8 ± 6,4 %) v porovnaní s vehikulom (100,0 ± 9,1 %, jednosmerná ANOVA, p < 0,05, priemer ± SEM). Expresia HER2 sa významne nelíšila vo všetkých liečebných skupinách (jednosmerná ANOVA, p = 0,1536). V prípade p-EGFR (Tyr1068) bola fosforylácia znížená na výrazne nižšiu úroveň kombinovanou liečbou melittínom a docetaxelom (9,0 ± 2,4 %) v porovnaní s vehikulom (100,0 ± 8,1 %, jednosmerná ANOVA, p < 0,0001, priemer SEM). Hladiny p-HER2 (Tyr1248) boli tiež znížené na významne nižšiu úroveň pri kombinovanej liečbe melittínom a docetaxelom (50,3 ± 7,8 %) v porovnaní s vehikulom (100,0 ± 5,6 %, jednosmerná ANOVA, p < 0,0001, priemer ± 0,0001, SEM). Pokles fosforylácie EGFR a HER2 in vivo po liečbe melitínom je v súlade s pozorovanými účinkami melitínu pri znižovaní fosforylácie týchto RTK v bunkách SKBR3, SUM159 a MDA-MB-231 (obr. 4a; doplnkový obrázok 4).
Diskusia
Apiterapia je novovznikajúca oblasť s potenciálom globálne ovplyvniť ekonomické aspekty výskumu rakoviny, najmä v komunitách s nedostatočnými zdrojmi. K dnešnému dňu však štúdie ešte musia úplne preskúmať molekulárny mechanizmus účinku včelieho jedu a melitínu a ich následné optimálne využitie v onkologickej aréne sa ešte musí komplexne preskúmať, najmä na liečbu rakoviny prsníka, ktorá sa najčastejšie vyskytuje. rakovina u žien na celom svete2. TNBC a nádory obohatené o HER2 sú vysoko agresívne podtypy rakoviny prsníka. TNBC je spojená s najvyššou mortalitou a napriek častej expresii EGFR bežne vykazuje rezistenciu na anti-EGFR terapie s vysokou závislosťou od signalizácie PI3K/Akt pre proliferáciu, prežitie a rezistenciu na chemoterapiu34.
Terapia anti-HER2 podstatne zlepšila dlhodobé prežívanie v skorých štádiách HER2-pozitívnych rakovín, ale u väčšiny pacientov v neskorom štádiu sa nakoniec vyvinula rezistencia a podľahli ochoreniu33,35,36. Nielenže sme preukázali selektivitu včelieho jedu a melitínu pre malígne bunky, ale odhalili sme aj vyššiu potenciu pre tieto agresívne typy rakoviny prsníka.
Tu ukazujeme, že včelí jed a melitín potláčajú ligandom indukovanú fosforyláciu EGFR a HER2, čím dynamicky modulujú downstream signálne dráhy v bunkách rakoviny prsníka. Navrhujeme, aby melitín priamo alebo nepriamo inhiboval dimerizáciu RTK. Melitín môže tiež vstúpiť do bunky, aby priamo alebo nepriamo moduloval downstream signálne dráhy25,60. Predchádzajúca práca ukázala, že melitín môže byť zacielený na bunkové línie nadmerne exprimujúce HER2 pomocou imunolipozómov nesúcich trastuzumab61. Tu demonštrujeme, že samotný melitín sa selektívne zameriava na bunky rakoviny prsníka nadmerne exprimujúce HER2 a EGFR. Je zaujímavé, že melitín bol pre bunky rakoviny prsníka toxickejší v porovnaní s včelím jedom, čo si vyžaduje ďalšie skúmanie.
V našej štúdii sme sa zamerali na bunkové línie SUM159 a SKBR3. SUM159 je bunková línia TNBC, ktorá exprimuje génový produkt EGFR a obsahuje mutácie missense v PI3KCA (H1047L) a v HRAS (G12D)62,63. Naopak, SUM159 je KRAS, NRAS, BRAF, PTEN a MAP2K4 divokého typu a negatívny na aktiváciu AKT1 a amplifikáciu AKT2 a AKT363. SKBR3 je HER2-obohatená bunková línia rakoviny prsníka, ktorá nadmerne exprimuje HER2 génový produkt64 a je KRAS, HRAS, NRAS, BRAF, PTEN, PI3KCA a MAP2K4 divokého typu63,65,66 a tiež negatívna na aktiváciu AKT1 a AKT2 a AKT3 zosilnenie63. Berúc do úvahy tieto molekulárne charakteristiky, downstream signálne dráhy EGFR nie sú konštitutívne aktivované v bunkách SUM159, napriek existujúcim mutáciám v HRAS a PI3KCA, pretože nie sú dostatočné na bazálnu aktiváciu týchto dráh67.
Uvádzame silnú a synergickú protinádorovú odpoveď s melitínom a docetaxelom vo vysoko agresívnom modeli TNBC in vivo. To zdôrazňuje potenciál melittínu na použitie v kombinovaných terapiách na potenciálne zvýšenie účinnosti a/alebo zníženie dávky cytotoxických činidiel, čo umožňuje dodanie nákladovo efektívnejšej liečby s potenciálne menšími vedľajšími účinkami. Melittin tiež znížil hladiny proteínu PD-L1 imunitného kontrolného bodu, ktorý sa podieľa na vyhýbaní sa imunite. Melittín by teda mohol znížiť imunosupresívne účinky nádorového mikroprostredia, ktoré prevládajú v TNBC v prítomnosti chemoterapie. To pridáva k údajom z predchádzajúcich správ, ktoré ukazujú, že melitín môže tiež znížiť populáciu makrofágov súvisiacich s nádorom typu M2 v mikroprostredí nádoru v modeli karcinómu pľúc68. Predpokladáme, že v našom in vivo modeli T11 môže signalizácia EGFR a HER2 modulovať expresiu PD-L1 v nádorových bunkách. Podľa predchádzajúcich imunohistochemických štúdií má PD-L1 najvyššiu expresiu v nádoroch TNBC, po ktorých nasledujú nádory obohatené o HER269,70,71,72 a expresia PD-L1 je spojená so slabým prežitím69. Pri bazálnych rakovinách prsníka sa ukázalo, že neprítomnosť proteínu ALIX koreluje s aktiváciou EGFR, čo zhoršuje biogenézu exozómov73. PD-L1 sa vylučuje prostredníctvom exozómov spôsobom závislým od ALIX, takže poškodenie exozómov zvyšuje PD-L1 na bunkovej membráne. Downregulácia ALIX podporuje prežitie nádoru zvýšením aktivácie EGFR a akumuláciou membrány PD-L1, čo vedie k imunosupresii73. Pri rakovine prsníka obohatenej o HER2 je presluch medzi HER2 a PD-L1 nedostatočne pochopený74. Avšak v HER2-pozitívnych bunkách rakoviny prsníka kokultivovaných s ľudskými mononukleárnymi bunkami periférnej krvi a na myšom modeli viedla terapia trastuzumabom proti HER2 k upregulácii PD-L175,76. Preto by inkorporácia melittínu s trastuzumabom mohla zrušiť túto imunosupresívnu reakciu.
Selektivita melitínu pre nádory riadené HER2 tiež predstavuje ďalší dôvod na kombináciu s látkami zameranými na HER2, vrátane monoklonálnych protilátok, trastuzumab-emtansínu a iných konjugátov protilátka-liečivo, kde by vlastnosti melitínu narušujúce membrány mohli zvýšiť kinetiku internalizácie cytotoxické zaťaženie. Naša práca tiež odhaľuje nové príležitosti na modifikáciu špecifických oblastí melitínu, aby sa ďalej zvýšila účinnosť a cielená špecifickosť pre malígne bunky. Upravené cielené peptidy, ako je RGD1-melittín, by sa mohli podávať intravenózne, aby sa umožnilo selektívnejšie navádzanie a príjem do nádorových buniek. Melittín by sa mohol dodávať aj prostredníctvom cielených nanočasticových prístupov, ako sú tie, ktoré boli predtým hlásené u „nanovčiel“77,78. Mohlo by sa tiež využiť prepojenie melitínu s toxínmi alebo proliečivami, ako sa uvádza pri fúziách melittínu štiepiteľných uPA79. Pred pokusmi na ľuďoch budú potrebné budúce štúdie na formálne posúdenie toxicity a maximálnych tolerovaných dávok týchto peptidov.
Včelí jed je celosvetovo dostupný a ponúka nákladovo efektívne a ľahko dostupné možnosti liečby v odľahlých alebo menej rozvinutých regiónoch. Bude potrebný ďalší výskum na posúdenie, či jed niektorých genotypov včiel má silnejšie alebo špecifickejšie protirakovinové aktivity, ktoré by sa potom dali využiť. Okrem rakoviny prsníka nádory nadmerne exprimujúce EGFR zahŕňajú rakovinu pľúc, glioblastóm a kolorektálny karcinóm80 a nádory, ktoré môžu nadmerne exprimovať HER2, zahŕňajú rakovinu žalúdka, vaječníkov, endometria, močového mechúra, pľúc, hrubého čreva a hlavy a krku81. Celkovo by sa naše výsledky mohli využiť na pomoc pri vývoji nových terapeutických modalít pre mnohé typy rakoviny spojené s častou rezistenciou na lieky a zlou prognózou.
Metódy
Chemické činidlá a protilátky
Všetky peptidy boli zakúpené od China Peptides Corporation, Ltd. Fluorescenčná fluoresceínová izotiokyanátová značka (FITC) bola konjugovaná na N-koniec FITC-melitín, SV40-melitín, TAT-melitín a EN1-mutant. CellTiter-Glo 2.0 z testu Luminescent Cell Viability Assay, NanoLuc-EGFR, FuGENE a furimazín boli všetky získané od Promega. TAMRA-EGF bol získaný od Invitrogen (Thermo Fisher Scientific). Docetaxel (kat. č. D-1000) bol získaný od LC Laboratories. Monoklonálna protilátka proti a-tubulínu (1:5000, kat. č. T5168), Hoechst (1:5000, kat. č. 94403) a ľudský EGF (kat. č. E9644) boli získané od Sigma-Aldrich. Myší EGF (kat. č. 315-09) bol získaný od Peprotech. Protilátky proti fosfo-HER2 (Tyr1248) (imunoblotting: 1:1000, imunohistochémia: 1:100, kat. č. 2247), fosfo-EGFR (Tyr1068) (imunoblotting: 1:1000, kat. č. 2234; imunohistochémia :350, kat. č. 3777, klon D7A5), fosfo-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) (1:2000, kat. č. 4370), fosfo-Akt (Ser473) (1: 2000, kat. č. 4060), fosfo-Akt (Thr308) (1:1000, kat. č. 13038), fosfo-SAPK/JNK (Thr183/Tyr185) (1:1000, kat. č. 4668), fosfo -p38 MAPK (Thr180/Tyr182) (1:1000, kat. č. 4511), celkový AKT (1:1000, kat. č. 9272 a 4685), štiepená kaspáza-3 (Asp175) (1:1000, kat. č. č. 9661), Ki-67 (1:400, kat. č. 9449), vzorkovacia súprava inhibítorov protilátky Jak/Stat Pathway (1:1000, kat. č. 8343) a sekundárny anti-myší IgG, HRP -protilátka (1:10 000, kat. č. 7076) a anti-králičí IgG, protilátka spojená s HRP (1:10 000, kat. č. 7074) boli vyrobené spoločnosťou Cell Signaling Technology. Monoklonálne protilátky proti ErbB2 (imunoblotting: 1:1000, imunohistochémia: 1:100, kat. č. ab8054, klon CB11), EGFR (imunoblotting: 1:5000, imunohistochémia: 1:100, kat. č. ab52894, klon a PD-L1 [PD-L1/2746] (1:100, kat. č. ab238697) boli vyrobené spoločnosťou Abcam. Sekundárne protilátky Alexa Fluor 488 kozia anti-myšia (1:500, kat. č. A11001) a Alexa Fluor 594 kozia anti-králičia (1:500, kat. č. A11012) boli získané od Thermo Fisher Scientific. Polyklonálna kozia anti-myšia IgG špecifická pre y-reťazec sekundárna protilátka (ELISA: 1:1000, kat. č. AP503P) bola získaná od Millipore. Bola vyrobená myšacia monoklonálna IgG protilátka špecifická pre ľudský IL-12 (28/00 8C1-6) použitá ako kontrolná protilátka pre experimenty ELISA a myšacia monoklonálna IgG protilátka špecifická pre melitín (ELISA: 1:350, klon 3B9). v zariadení na výrobu monoklonálnych protilátok v Ústave lekárskeho výskumu Harryho Perkinsa. Test TUNEL (In situ Cell Death Detection Kit) bol získaný od Roche.
Zber včelieho jedu
Jed bol zozbieraný pomocou robotníc alebo kráľovien z niekoľkých rôznych populácií včiel Apid. Vzorky jedu odobraté od európskych včiel medonosných (Apis mellifera) a čmeliakov s chvostom (Bombus terrestris audax) pochádzajú z Perthu (Austrália), Dublinu (Írsko) a Londýna (Anglicko). Včelí jed bol odobratý od 30 robotníkov z každej z troch rôznych kolónií z včelína alebo farmy, ako je opísané. Včelí jed z Austrálie bol zozbieraný zo včelína spravovaného Centrom pre integrovaný výskum včiel (CIBER), ktorý sa nachádza na University of Western Australia (UWA: -31.980151, 115.817919). Včelí jed z Írska bol zozbieraný z jednej kolónie na včelíne na Trinity College Dublin (53,343933, −6,254635) a ďalších dvoch kolónií z fariem v blízkosti Glasnevin (53,383245, −6,276333) a Blanchardstown (50,73.584). Jed včiel a čmeliakov z Anglicka bol zozbieraný na Royal Holloway University of London (51,425626, −0,562987). Jed čmeliakov sa odobral od 20 robotníc z každej z 2 komerčne zakúpených kolónií, pričom jedna čmeliaková kráľovná z každej z týchto dvoch kolónií sa použila na odber jedu čmeliakových kráľovien. Nezávislé biologické hlavné zmesi sa pripravili tak, že sa jed z rôznych kolónií držal oddelene, pričom sa celkovo zhromaždil jed 312 včiel.
Žľazový jed sa odobral manuálnou disekciou. Včely boli odchytené v blízkosti vchodu do úľa pre včely alebo priamo z kolónie pre čmeliaky a anestetizované oxidom uhličitým a ochladené na ľade. Z každého jedinca bol vypreparovaný bodací aparát; potom sa odstránila jedová žľaza a umiestnila sa do fosfátom pufrovaného fyziologického roztoku (PBS). Žľazy boli prepichnuté ihlou Terumo (25 G × 5/8) a odstredené (13 000 g, 10 min, 4 °C) a supernatant bol odobratý, obsahujúci jed v kvapalnej suspenzii. Koncentrácia proteínu každej hlavnej zmesi bola kvantifikovaná pomocou proteínového testu kompatibilného s detergentom (Bio-Rad), meraním absorbancie pri 750 nm pomocou Millennium Science BioTek PowerWave XS2 (Gen 5 1.11 Software, Verzia 1.11.5). Každá hlavná zmes sa potom rozdelila na alikvóty a skladovala pri -80 °C.
Bunkové línie a kultivačné podmienky
Všetky bunkové línie boli zakúpené od American Type Culture Collection (Manassas, VA, USA), okrem buniek HEK293FT, ktoré boli zakúpené od Invitrogen (Thermo Fisher Scientific, Victoria, Austrália), SUM149 a SUM159, ktoré boli získané od Asterand Bioscience (Detroit, MI, USA) a bunky T11 a B.15, ktoré láskavo poskytli Charles Perou a Lyuba Varticovski z University of North Carolina v Chapel Hill a National Institutes of Health. T11 a B.15 sú veľmi dobre charakterizované bunkové línie37,38.
Bunky boli inkubované pri 37 ° C a 5 % CO2 a doplnené 1 % antibiotikom-antimykotikom. HDFa (normálne primárne dospelé ľudské dermálne fibroblasty) bunky sa kultivovali v DMEM s 10 % fetálnym bovinným sérom (FBS). MCF 10A a MCF-12A (imortalizované epitelové bunky ľudskej mliečnej žľazy, netransformované) sa udržiavali v DMEM/F-12 s doplnkami (5 % fetálne konské sérum, 20 ng/ml epidermálny rastový faktor, 10 μg/μl inzulín, 100 ng/ml toxín cholery a 500 ng/ml hydrokortizónu). Bunky NIH/3T3 (myšie embryonálne fibroblasty) sa udržiavali v DMEM s 10 % FBS. HEK293FT (bunky 293 ľudských embryonálnych obličiek stabilne exprimujúce SV40 veľký T antigén) sa kultivoval v DMEM s 10 % FBS a doplnkami (1 % glutamín a 0,4 mg/ml G418 Geneticin, Gibco). MCF7 (ľudský luminálny A karcinóm prsníka) sa udržiaval v MEM a s 10 % FBS a doplnkami (po 1 % pyruvát sodný, hydrogenuhličitan sodný a neesenciálne aminokyseliny). T-47D a ZR-75-1 (obe ľudské luminálna A rakovina prsníka) sa kultivovali v RPMI s 10 % FBS. MDA-MB-231 (ľudský karcinóm prsníka s nízkym obsahom klaudínu) sa kultivoval v DMEM s 10 % FBS. SUM149 (ľudský bazálny karcinóm prsníka) bol kultivovaný v F-12 s 10 % FBS. SUM159 (ľudský karcinóm prsníka s nízkym obsahom klaudínu) bol kultivovaný v F-12 s 5% FBS a doplnkami (5 μg/ml inzulínu a 1 μg/ml hydrokortizónu). MDA-MB-453 (rakovina prsníka obohatená o ľudský HER2) sa kultivovala v DMEM s 10 % FBS. SKBR3 (rakovina prsníka obohatená o ľudský HER2) sa kultivovala v RPMI s 10 % FBS a 1 % pyruvátom sodným. p53-T11 (myší karcinóm prsníka s nízkym obsahom klaudínu) sa udržiaval v médiu RPMI 1640 s 10 % FBS. BRCA− B.15 (myší bazálny karcinóm prsníka) sa udržiaval v médiu RPMI 1640 s 10 % FBS.
Testy bunkovej životaschopnosti
Životaschopnosť buniek bola stanovená luminiscenčným testom životaschopnosti buniek podľa protokolu dodávateľa. Bunky boli nanesené na 96-jamkové kultivačné platne a inkubované pri 37 °C a 5 % C02 počas 24 hodín. Pre testy odozvy na dávku sa médium zlikvidovalo a nahradilo médiom obsahujúcim uvedené koncentrácie včelieho jedu alebo peptidu a kultivovalo sa 24 hodín. Pre životaschopnosť buniek počas 60 minút boli bunky ošetrené IC50 včelieho jedu alebo melitínu pre každú bunkovú líniu v krátkych časových intervaloch počas 1 hodiny a životaschopnosť bola stanovená ihneď po ošetrení. Na stanovenie životaschopnosti sa bunky inkubovali s činidlom CellTiter-Glo (CTG) 2.0 počas 10 minút. Životaschopnosť buniek sa kvantifikovala meraním luminiscencie pomocou čítačky EnVision 2102 Multilabel Reader (PerkinElmer). Experimenty sa uskutočňovali v biologických replikátoch (n = 3).
Produkcia primárnej monoklonálnej protilátky proti melitínu
Produkcia protilátok sa uskutočňovala v súlade s protokolmi schválenými Výborom pre etiku zvierat z Ústavu lekárskeho výskumu Harryho Perkinsa. Samice myší A/J boli imunizované včelím jedom zozbieraným v Austrálii. Myši dostali intraperitoneálne injekcie 12 μg jedu v kompletnom Freundovom adjuvans (Difco), po ktorých nasledovala posilňovacia dávka nekompletného Freundovho adjuvans na 29. deň a vodná posilňovacia dávka PBS v dávke 7 μg/myš na 49. deň. a séra boli testované pomocou ELISA. Najlepšia odpoveď bola posilnená 7 μg včelieho jedu v PBS 4 dni pred fúziou. Slezinové bunky boli fúzované s Sp2/O myelómovými bunkami podľa štandardných postupov82. Supernatanty obsahujúce protilátky sa testovali pomocou ELISA. Hybridómový klon 3B9 bol vybraný na ďalšiu štúdiu. Protilátka bola produkovaná pestovaním hybridómových buniek v bioreaktoroch v Hybridoma Serum Free Medium (Gibco). Protilátka bola purifikovaná pomocou proteín G-Sepharose chromatografie. Purifikovaná protilátka bola dialyzovaná v PBS (pH 7,3). Protilátka bola odteraz označovaná ako anti-melitínová protilátka (3B9).
Enzymovo viazaný imunosorbentový test (ELISA)
Jedy a peptidy boli nanesené na 96-jamkové doštičky s čírou krivkou pri 5 μg/ml v uhličitanovom pufri a inkubované pri 4 °C počas 24 hodín. Kvapalina sa odstránila a doštičky sa trikrát premyli v roztoku 0,05 % TWEEN-20 („Tween-20,“ Sigma-Aldrich) v PBS. Primárne protilátky sa pridali do jamiek s riedením 1:2 počínajúc od 10 μg/ml v riedidle (0,1 % bovinný sérový albumín (BSA) v PBS) a inkubovali sa 1 hodinu pri teplote miestnosti. Primárne protilátky sa odstránili a doštičky sa trikrát premyli v 0,05 % Tween-20 v PBS. Do jamiek sa pridala polyklonálna kozia anti-myšia IgG špecifická pre y-reťazec sekundárna protilátka (1:1000 v riedidle) a inkubovala sa 1 hodinu pri teplote miestnosti. Primárne protilátky sa odstránili a doštičky sa trikrát premyli v 0,05 % Tween-20 v PBS. Do jamiek sa pridal ELISA-vyvíjací pufor, roztok čistenej vody obsahujúcej 10 % kyseliny citrónovej (pH 4,2), 2 % ABTS a 0,1 % H202, a platne sa inkubovali v tme pri teplote miestnosti 15 minút. Absorbancia sa zaznamenávala pri 405 nm s použitím čítačky doštičiek VICTOR Light so softvérom Wallac 1420 Manager (PerkinElmer). Kontrolou bola myšacia monoklonálna IgG protilátka (28/00 8C1-6), ktorá reaguje s ľudským IL-12, aplikovaná na melitínový peptid na doštičke ELISA. Experimenty sa uskutočňovali v biologických replikátoch (n = 3).
Kompetičné experimenty anti-melittínových protilátok
HDFa a SUM159 bunky boli nanesené na 96-jamkové kultivačné platne a inkubované pri 37 °C a 5 % C02 počas 24 hodín. Zvyšujúce sa koncentrácie anti-melitínovej protilátky sa inkubovali s IC50 koncentráciami včelieho jedu alebo melitínu pre každú bunkovú líniu počas 1 hodiny pri teplote miestnosti a potom sa pridali k bunkám na 24 hodín. Životaschopnosť buniek bola stanovená tak, ako je opísané v časti „Skúšky životaschopnosti buniek“. Experimenty sa uskutočňovali v biologických replikátoch (n = 3).
Western blot
Bunky boli nanesené na 6-jamkové platne v hustote 300 000 buniek/jamka a inkubované pri 37 °C a 5 % C02 počas 24 hodín. Experimenty s bunkovou kultúrou sa uskutočňovali tak, ako je opísané, a potom sa postupovalo podľa štandardného protokolu Western blot, ako je tu opísané. Bunky boli premyté studeným PBS a lyzované studeným proteínovým lyzačným pufrom (2 % dodecylsulfát sodný (SDS), 125-mmol/l Tris-HCl, pH 6,8). Vzorky sa sonikovali počas 10 s pri 10 mA a koncentrácie proteínov sa kvantifikovali pomocou testu proteínov kompatibilných s detergentom (Bio-Rad). Rovnaké množstvá proteínov sa zmiešali s nanášacím pufrom (Laemmli Sample Buffer, Bio-Rad) doplneným o redukčné činidlo ditiotreitol (DTT). Vzorky bielkovín boli denaturované varom pri 95 °C počas 5 minút, vložené do prefabrikovaných gélov Mini-PROTEAN (Bio-Rad) a podrobené elektroforéze pri 100 V a potom prenesené na PVDF membrány (Bio-Rad) pomocou Trans-Blot Turbo Transfer System (Bio-Rad) na 7 min. Membrány sa inkubovali s TBST (20 mM Tris-HCl, pH 7,4, 150 mM NaCI a 0,1 % Tween-20) s 5 % odtučneným mliekom, aby sa zablokovala nešpecifická väzba. Membrány sa inkubovali cez noc pri 4 °C s primárnymi protilátkami zriedenými v 3 % BSA a 0,02 % azide sodnom. Signál bol detegovaný pomocou Luminata Crescendo Western HRP Substrate (Millipore) s ChemiDoc MP Imaging System (Bio-Rad) so softvérom Image Lab Software (Bio-Rad, verzia 6). Western bloty boli odvodené z rovnakého experimentu a spracované paralelne. Neorezané skeny Western blotov sú uvedené na doplnkových obr. 10-15.
Prietoková cytometria
Apoptóza a nekróza boli hodnotené pomocou Annexin V-FITC Apoptosis Detection Kit I (BD Biosciences) podľa protokolu výrobcu. Bunky SUM159 boli nanesené na 6-jamkové kultivačné platne počas 24 hodín. Médium sa potom vyhodilo a nahradilo médiom obsahujúcim včelí jed alebo melittín (koncentrácie IC50) a kultivovalo sa 60 minút. Bunky boli zhromaždené s trypsínom a médiom a centrifugované (1000 g, 5 min, 24°C), premyté studeným PBS, centrifugované (1000g, 5 min, 24°C) a resuspendované v 1x väzbovom pufri. Bunky boli pripravené na koncentráciu 1 milión buniek/ml v 1x väzbovom pufri. Vzorky boli inkubované s FITC a PI (5 ul každého) v tme počas 15 min. Prítomnosť živých, mŕtvych, apoptotických alebo nekrotických buniek bola hodnotená pomocou BD Accuri C6 Cytometer (BD Biosciences, San Jose, USA) so softvérom BD Accuri C6 a analyzovaná pomocou FlowJo™ (Ashland, USA, Windows Verzia 7). Experimenty sa uskutočňovali v biologických replikátoch (n = 3). Stratégie hradlovania sú uvedené na doplnkovom obrázku 16.
Mikroskopia živých buniek
Bunky SKBR3 boli nanesené do mikrojamkovej misky so skleneným dnom (10 x 35 mm, MatTek) a inkubované počas 24 h. Miska s mikrojamkami sa nechala ekvilibrovať v inkubačnej komore na vrchnej časti stolíka konfokálneho mikroskopu NIKON Eclipse Ti (37 °C a 5 % CO2) počas 20 minút. Objektív 20x sa použil s Kohlerovým zarovnaním a snímky sa snímali každú minútu od 10 minút pred do 1 hodiny po ošetrení s IC50 včelieho jedu zozbieraného v Austrálii. Autori oceňujú zariadenia a vedeckú a technickú pomoc, ktorú ponúka National Imaging Facility, schopnosť Národnej stratégie pre kolaboratívnu výskumnú infraštruktúru (NCRIS), ako aj Austrálske výskumné zariadenie pre mikroskopiu a mikroanalýzu, obe v Centre pre mikroskopiu, charakterizáciu a analýzu ( CMCA), UWA, zariadenie financované univerzitami, vládami štátov a Commonwealthu.
Skenovacia elektrónová mikroskopia
Sklenené krycie sklíčka (priemer 12 mm, Menzel, Thermo Fisher Scientific) boli potiahnuté hydrobromidom poly-L-lyzínu (Sigma-Aldrich) počas 20 minút a potom dvakrát premyté čistenou vodou. Bunky SUM159 boli nanesené na podložné sklíčka v hustote 62 500 buniek/jamka a inkubované pri 37 °C a 5 % C02 počas 24 hodín. Bunky boli dvakrát premyté PBS a potom ošetrené vehikulom alebo IC50 koncentráciami včelieho jedu a melitínu počas 1 hodiny. Bunky sa premyli dvakrát PBS, potom sa fixovali 4% formaldehydom v PBS počas 25 minút a potom sa znova trikrát premyli PBS. Pri príprave na mikroskopiu boli vzorky ponorené do 2,5 % glutaraldehydu a inkubované pri 4 °C počas 2 hodín. Vzorky boli premyté deionizovanou vodou a ponorené do zvyšujúcich sa koncentrácií etanolu (50 %, 70 %, 95 %, 100 % a potom 100 % absolútny „suchý“ etanol). Medzi každým ponorením boli vzorky dehydratované v špecializovanej mikrovlnnej rúre (PELCO, BioWave 34700 Laboratory Microwave System). Proces dehydratácie bol dokončený pomocou zariadenia na sušenie kritického bodu E3000, aby sa etanol vo vzorke nahradil superkritickým CO2. Spracované krycie sklíčka boli namontované na držiaky SEM (ProSciTech) s uhlíkovými plôškami. Vzorky boli potiahnuté 3 nm platinou, aby boli elektronicky vodivé pred vizualizáciou pod skenovacím elektrónovým mikroskopom (Zeiss 1555 VP-FESEM) v CMCA, UWA. Snímky boli urobené detektorom v šošovke pri pracovnej vzdialenosti 2,6 mm, clone 30 μm a urýchľovacím napätí 5 kV. Obrázky sa analyzovali pomocou softvéru na analýzu obrazu FIJI (ImageJ)83.
Imunofluorescencia
Sklenené krycie sklíčka (priemer 12 mm, Menzel, Thermo Fisher Scientific) sa umiestnili do 24-jamkových doštičiek a potiahli sa poly-L-lyzínom (Sigma-Aldrich) počas 20 minút a potom sa dvakrát premyli čistenou vodou. Bunky SUM159 boli nanesené na podložné sklíčka a inkubované pri 37 °C a 5 % C02 počas 24 hodín. Bunky boli ošetrené počas 30 minút vehikulom alebo IC50 včelieho jedu, melitínu, RGD1-melitínu a ekvivalentnej molárnej koncentrácie ako melitínu pre DEDE-melittín. Bunky sa premyli dvakrát PBS, potom sa fixovali 4% paraformaldehydom v PBS počas 25 minút a potom sa znova trikrát premyli PBS. Väzba nešpecifickej protilátky bola blokovaná použitím 5% normálneho kozieho séra (Thermo Fisher Scientific) v PBS počas 1 hodiny pri teplote miestnosti. K bunkám boli pridané primárne protilátky, vrátane monoklonálnej protilátky proti melittínu (5 ug/ml) a 1:500 anti-EGFR EP38Y. Vzorky sa inkubovali za jemného kývania pri 4 °C cez noc. Bunky sa trikrát premyli PBS a potom sa inkubovali s 1:500 kozej anti-myšej sekundárnej protilátky Alexa Fluor 488, 1:500 kozej anti-králičej sekundárnej protilátky Alexa Fluor 594 a Hoechst (1:5000) v PBS pri izbovej teplote počas 1 hodiny. Vzorky boli trikrát premyté PBS a pripevnené na sklenené krycie sklíčka pomocou SlowFade Diamond Antifade Mountant (Thermo Fisher Scientific). Sklíčka sa zobrazili pomocou konfokálneho fluorescenčného invertovaného mikroskopu Nikon Ti-E. Snímky boli urobené pomocou 20x vzduchového objektívu (NA 0,75) a sekvenčnej excitácie s použitím vlnových dĺžok 405 nm (Hoechst 34580), 488 nm (sekundárna protilátka Alexa Fluor 488) a 561 nm (sekundárna protilátka Alexa Fluor 594). Obrázky boli zozbierané pomocou softvéru NIS-C Elements Software a spracované pomocou FIJI (ImageJ) na CMCA83.
Bioluminiscenčný rezonančný prenos energie (BRET)
Interakcie receptor-ligand sa hodnotili pomocou BRET s použitím metódy podobnej tej, ktorá bola opísaná vyššie84,85. BRET zahŕňa nežiarivý prenos energie (dipól-dipól) medzi dvoma proteínmi alebo molekulami, ktoré sú predmetom záujmu, označených buď donorovou luciferázou alebo akceptorovým fluorofórom po oxidácii substrátu luciferázou a následnej emisii svetla54. FITC značky boli konjugované na N-koniec melitínu (FITC-melitín) a DEDE-melittín (FITC-DEDE-melitín). Bunky HEK293 stabilne exprimujúce SV40 veľký T antigén (HEK293FT) sa naniesli na 6-jamkové platne v hustote 550 000 buniek/jamku počas 24 hodín. Bunky HEK293FT boli transfekované plazmidmi obsahujúcimi cDNA pre NanoLuc-EGFR s použitím FuGENE. Stručne, plazmidová cDNA sa inkubovala 10 minút pri teplote miestnosti so zmesou transfekčného činidla a DMEM bez séra v pomere 10 ng/ul NanoLuc-EGFR: 4 μl FuGENE: 100 μl SFM. Zmes sa pridala k bunkám HEK293FT v konečnej koncentrácii 10 ng/ul NanoLuc-EGFR na jamku 6-jamkovej platne a bunky sa inkubovali 24 h. Bunky sa premyli PBS a oddelili sa trypsínom, potom sa zhromaždili v médiu obsahujúcom 5 % fetálneho teľacieho séra v DMEM bez fenolovej červene. Bunky boli nanesené v množstve 50 000 buniek/jamku na 96-jamkové biele platne potiahnuté poly-L-lyzínom a inkubované počas 24 hodín. Pre saturačné aj kinetické testy BRET sa použili dva filtre na súčasné meranie luminiscencie s krátkou a dlhou vlnovou dĺžkou zodpovedajúcou emisným vlnovým dĺžkam donorových a akceptorových molekúl.
Pre experimenty s kinetikou asociácie ligandov v reálnom čase sa médium odstránilo z buniek, ktoré sa potom inkubovali s 50 μl/jamku NanoLuc substrátu furimazínu do konečnej koncentrácie 10 μM zriedeného v Hankovom vyváženom soľnom roztoku (HBSS). Bunky sa potom ekvilibrovali v čítačke platní CLARIOstar (BMG Labtech, Austrália) počas 5 minút, aby sa zaznamenali bazálne hodnoty. Ligandy (TAMRA-EGF, FITC-melitín a FITC-DEDE-melittín) sa potom pridali do rozsahu správnych konečných koncentrácií a záznamy NanoBRET sa robili každých 90 s počas 60 minút pri 37 °C. Pre saturačné experimenty sa médium z buniek odstránilo a pridal sa rozsah koncentrácií TAMRA-EGF, FITC-melittín a FITC-DEDE-melittín v prítomnosti alebo neprítomnosti konkurenčnej koncentrácie (1 uM) neznačeného EGF. a inkubované pri 37 °C počas 60 minút v tme. Furimazín sa pridal v konečnej koncentrácii 10-uM. Nahrávky boli urobené pomocou LUMIstar Omega (BMG Labtech, Austrália). Údaje sú prezentované ako „surový pomer BRET“, odvodený z pomeru dlhovlnnej emisie (akceptor) k emisii s krátkou vlnovou dĺžkou (donora). Experimenty sa uskutočňovali v biologických replikátoch (n = 3).
Analýza kombinovaných účinkov liekov
Včelí jed alebo melitín boli kombinované s docetaxelom a podávané v koncentráciách uvedených v nekonštantnom pomere v bunkách T11 počas 24 hodín. Životaschopnosť buniek sa hodnotila pomocou CellTiter-Glo, ako bolo uvedené vyššie. Kombinovaný účinok včelieho jedu alebo melittínu s docetaxelom sa hodnotil metódou mediánu dávka-účinok pomocou softvéru CompuSyn (ComboSyn). Táto metóda určuje CI na základe účinku kombinácie dvoch látok (kde CI < 1 je synergický, CI > 1 je antagonistický a CI = 1 je aditívny)56. Experimenty sa uskutočňovali v biologických replikátoch (n = 3).
Zvierací model a liečba
Tieto pokusy na zvieratách sa uskutočnili v súlade s protokolmi schválenými Výborom pre etiku zvierat UWA. Na simuláciu pokročilého modelu rakoviny prsníka s nízkym obsahom claudínu sa 2,5 × 105 buniek T11 suspendovalo v médiu bez séra a BD Matrigel Matrix High Concentration (BD Bioscience) v pomere 1:1 na celkový objem 100 μl a injikovalo sa subkutánne. do bokov 5-týždňových samíc BALB/cJ (Animal Resources Centre, WA, Austrália) pomocou 26-G ihly. Použité bunky T11 boli lentivírusovo transdukované konštruktom ZsGreen-luciferáza a trikrát triedené, aby sa dosiahlo obohatenie lepšie ako 99 % buniek pozitívnych na luciferázu. Melittín sa suspendoval v Milli-Q voda + 5 % dextróza. Docetaxel (v prášku) sa suspendoval v 25 % TWEEN 80 (Sigma-Aldrich) a 75 % zmesi 15,25:84,75 (v/v) roztoku absolútneho etanolu a čistenej vody a udržiaval sa pri -20 °C. Bezprostredne pred ošetrením bol docetaxel čerstvo zriedený v Milli-Q voda + 5 % dextróza na požadovanú konečnú koncentráciu. Tri dni po vytvorení nádorov T11 (~50 mm3) boli myši náhodne rozdelené do 4 skupín (n = 12 myší/skupina). Liečby sa injikovali intratumorálne v dňoch 3, 5, 7, 9, 11, 13 a 15 po inokulácii buniek T11 vehikulom, melitínom (5 mg/kg), docetaxelom (7 mg/kg) alebo kombináciou melitín (5 mg/kg) a docetaxel (7 mg/kg). Zvieratá sa monitorovali na veľkosť nádoru každé 2 dni a objemy sa vypočítali podľa modifikovaného elipsoidného vzorca (objem = = = šírka 2 x x dĺžka/2). Zvieratá boli humánne usmrtené, keď nádory dosiahli 800 mm3.
Imunohistochemická analýza nádorov
Nádorové tkanivá boli fixované v 4% paraformaldehyde, premyté trikrát v PBS a ponechané v 70% etanole. Nádory sa zaliali do parafínu a pripravili sa 5 μm rezy. Na farbenie hematoxylínom/eozínom boli sklíčka zbavené vosku, hydratované pomocou banky s klesajúcim roztokom etanolu, zafarbené Gillovým hematoxylínom, dehydratované pomocou 70 % etanolu, zafarbené eozínom, ďalej dehydratované pomocou 100 % etanolu, vyčistené pomocou toluénu a pripevnené na krycie sklíčka pomocou Montážne médium Acrymount IHC (StatLab). Apoptóza nádorových buniek bola stanovená v tkanivových rezoch testom TUNEL (In situ Cell Death Detection Kit, Roche).
Bioluminiscenčné zobrazovanie
Na presné sledovanie zmien v raste nádoru in vivo s liečbou sme vykonali bioluminiscenčnú analýzu pomocou zobrazovacieho systému Caliper IVIS Lumina II v CMCA, UWA. Analýzy sa uskutočňovali každé 2 dni po vytvorení nádorov. Myšiam sa intraperitoneálne injikovalo 200 ul D-Luciferínu (Cayman Chemical) vo finálnej koncentrácii 150 mg/kg rozpusteného v PBS pred anestetizáciou 4 % izofluránom. Po anestézii sa myši umiestnili do predhriatej komory bioluminiscenčného zobrazovača a zobrazili sa 7–12 minút po injekcii pod 2% izofluránom, kým intenzita bioluminiscenčného signálu nedosiahla ustálený stav.
Štatistická analýza
Všetky údaje boli odvodené z viacerých experimentov uskutočnených aspoň trojmo. Štatistické analýzy sa uskutočnili pomocou GraphPad Prism v8 (GraphPad Software Inc.), Office Excel 365 (Microsoft) a SPSS Predictive Analytics Software (IBM, verzia 26). Pre testy bunkovej životaschopnosti sa údaje normalizovali na priemernú luminiscenciu stavu vehikula, ktorá sa považovala za 100% životaschopnosť, s IC50 odvodenými v GraphPad Prism. Na imunohistochémiu v liečených nádoroch T11 na detekciu p-HER2 (Tyr1248) a p-EGFR (Tyr1068) sa vehikulum normalizovalo na 100 %. Kde je to vhodné a ako je uvedené v hlavnom texte, štatistická významnosť sa stanovila pomocou nepárových dvojstranných Studentových t testov, nepárovej jednosmernej ANOVA s Tukeyho post hoc testom HSD korigujúcim viacnásobné porovnania, obojsmernej ANOVA s opakovanými meraniami nasledovanými Sidakovým alebo Tukeyho viacnásobný porovnávací test alebo zovšeobecnený lineárny model (GLM). Pre všetky testy sa rozdiely považovali za významné pri p < 0,05 (), p < 0,01 () a p < 0,001 ().
Súhrn prehľadov
Ďalšie informácie o dizajne výskumu sú k dispozícii v súhrne správ o výskume prírody, ktorý je prepojený s týmto článkom.
Dostupnosť údajov
Všetky údaje generované alebo analyzované počas tejto štúdie sú zahrnuté v tomto publikovanom článku (a jeho doplnkových informačných súboroch). Protilátka proti melittínu vyvinutá v zariadení na výrobu monoklonálnych protilátok v Inštitúte lekárskeho výskumu Harryho Perkinsa by mohla byť sprístupnená po uzavretí príslušných dohôd.
História zmien
- september 2020Pôvodná verzia tohto článku bola po zverejnení aktualizovaná, pretože sa neúmyselne zmenili odkazy na súbory Súhrn prehľadov a Doplnkové informácie. Chyba s odkazmi bola opravená pre súbory Súhrn prehľadov a Doplnkové informácie vo verzii článku HTML.
Referencie
- Son, D. J. et al. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 115, 246–270 (2007).PubMed CAS Article Google Scholar
- Fitzmaurice, C. et al. Global, regional, and national cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life-years for 32 Cancer Groups, 1990 to 2015. JAMA Oncol. 3, 524 (2017).PubMed Google Scholar
- Park, S.-C. et al. Investigation of toroidal pore and oligomerization by melittin using transmission electron microscopy. Biochem. Biophys. Res. Commun. 343, 222–228 (2006).PubMed CAS Article Google Scholar
- Lyu, Y., Zhu, X., Xiang, N. & Narsimhan, G. Molecular Dynamics Study of Pore Formation by Melittin in a 1,2-Dioleoyl- sn -glycero-3-phosphocholine and 1,2-Di(9 Z -octadecenoyl)- sn -glycero-3-phospho-(1′- rac -glycerol) Mixed Lipid Bilayer. Ind. Eng. Chem. Res. 54, 10275–10283 (2015).CAS Article Google Scholar
- Terwilliger, T. C. & Eisenberg, D. The structure of melittin. II. Interpretation of the structure. J. Biol. Chem. 257, 6016–6022 (1982).PubMed CAS Article Google Scholar
- Lee, M.-T., Sun, T.-L., Hung, W.-C. & Huang, H. W. Process of inducing pores in membranes by melittin. Proc. Natl Acad. Sci. USA 110, 14243–14248 (2013).PubMed CAS Article PubMed Central Google Scholar
- Sun, D., Forsman, J. & Woodward, C. E. Multistep molecular dynamics simulations identify the highly cooperative activity of melittin in recognizing and stabilizing membrane pores. Langmuir 31, 9388–9401 (2015).PubMed CAS Article Google Scholar
- Tu, W. C., Wu, C. C., Hsieh, H. L., Chen, C. Y. & Hsu, S. L. Honeybee venom induces calcium-dependent but caspase-independent apoptotic cell death in human melanoma A2058 cells. Toxicon 52, 318–329 (2008).PubMed CAS Article Google Scholar
- Gao, D. et al. Melittin induces NSCLC apoptosis via inhibition of miR-183. Onco. Targets Ther. 11, 4511–4523 (2018).PubMed PubMed Central Article Google Scholar
- Sisakht, M. et al. Bee venom induces apoptosis and suppresses matrix metaloprotease-2 expression in human glioblastoma cells. Braz. J. Pharmacogn. 27, 324–328 (2017).CAS Article Google Scholar
- Killion, J. J. & Dunn, J. D. Differential cytolysis of murine spleen, bone-marrow and leukemia cells by melittin reveals differences in membrane topography. Biochem. Biophys. Res. Commun. 139, 222–227 (1986).PubMed CAS Article Google Scholar
- Jo, M. et al. Anti-cancer effect of bee venom toxin and melittin in ovarian cancer cells through induction of death receptors and inhibition of JAK2/STAT3 pathway. Toxicol. Appl. Pharmacol. 258, 72–81 (2012).PubMed CAS Article Google Scholar
- Zarrinnahad, H. et al. Apoptotic effect of melittin purified from iranian honey bee venom on human cervical cancer Hela cell line. Int. J. Pept. Res. Ther. 24, 563–570 (2018).PubMed CAS Article Google Scholar
- Wang, X. et al. Melittin-induced long non-coding RNA NONHSAT105177 inhibits proliferation and migration of pancreatic ductal adenocarcinoma. Cell Death Dis. 9, 940 (2018).PubMed PubMed Central Article CAS Google Scholar
- Zhu, H. G., Tayeh, I., Israel, L. & Castagna, M. Different susceptibility of lung cell lines to inhibitors of tumor promotion and inducers of differentiation. J. Biol. Regul. Homeost. Agents 5, 52–58 (1991).PubMed CAS Google Scholar
- Yu, X. et al. Immune modulation of liver sinusoidal endothelial cells by melittin nanoparticles suppresses liver metastasis. Nat. Commun. 10, 574 (2019).PubMed PubMed Central CAS Article Google Scholar
- Gajski, G. et al. Combined antitumor effects of bee venom and cisplatin on human cervical and laryngeal carcinoma cells and their drug resistant sublines. J. Appl. Toxicol. 34, 1332–1341 (2014).PubMed CAS Article Google Scholar
- Choi, K. et al. Cancer cell growth inhibitory effect of bee venom via increase of death receptor 3 expression and inactivation of NF-kappa B in NSCLC cells. Toxins 6, 2210–2228 (2014).PubMed PubMed Central Article CAS Google Scholar
- Shaw, P. et al. Synergistic effects of melittin and plasma treatment: a promising approach for cancer therapy. Cancers 11, 1109 (2019).PubMed Central CAS Article Google Scholar
- Ip, S. W. et al. The role of mitochondria in bee venom-induced apoptosis in human breast cancer MCF7 cells. Vivo 22, 237–245 (2008).Google Scholar
- Jeong, Y.-J. et al. Melittin suppresses EGF-induced cell motility and invasion by inhibiting PI3K/Akt/mTOR signaling pathway in breast cancer cells. Food Chem. Toxicol. 68, 218–225 (2014).PubMed CAS Article Google Scholar
- Jung, G. B. et al. Anti-cancer effect of bee venom on human MDA-MB-231 breast cancer cells using Raman spectroscopy. Biomed. Opt. Express 9, 5703 (2018).PubMed PubMed Central CAS Article Google Scholar
- Oršolić, N., Šver, L., Verstovšek, S., Terzić, S. & Bašić, I. Inhibition of mammary carcinoma cell proliferation in vitro and tumor growth in vivo by bee venom. Toxicon 41, 861–870 (2003).PubMed Article CAS Google Scholar
- Oršolić, N., Terzić, S., Šver, L. & Bašić, I. Honey-bee products in prevention and/or therapy of murine transplantable tumours. J. Sci. Food Agric. 85, 363–370 (2005).Article CAS Google Scholar
- Kohno, M., Horibe, T., Ohara, K., Ito, S. & Kawakami, K. The membrane-lytic peptides K8L9 and melittin enter cancer cells via receptor endocytosis following subcytotoxic exposure. Chem. Biol. 21, 1–11 (2014).Article CAS Google Scholar
- Lim, H., Baek, S. & Jung, H. Bee venom and its peptide component melittin suppress growth and migration of melanoma cells via inhibition of PI3K/AKT/mTOR and MAPK pathways. Molecules 24, 929 (2019).PubMed Central Article CAS Google Scholar
- Van Vaerenbergh, M., Debyser, G., Smagghe, G., Devreese, B. & De Graaf, D. C. Unraveling the venom proteome of the bumblebee (Bombus terrestris) by integrating a combinatorial peptide ligand library approach with FT-ICR MS. Toxicon 102, 81–88 (2015).PubMed Article CAS Google Scholar
- Qiu, Y. et al. Molecular cloning of the Bombus terrestris bumblebee venom protein phospholipase A 2 and its anti-leukemia effects on K562 cells. J. Asian Pac. Entomol. 20, 699–704 (2017).Article Google Scholar
- Prat, A. & Perou, C. M. Deconstructing the molecular portraits of breast cancer. Mol. Oncol. 5, 5–23 (2011).PubMed CAS Article Google Scholar
- Lehmann, B. D. et al. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 121, 2750–2767 (2011).PubMed CAS Article PubMed Central Google Scholar
- Mayer, I. A., Abramson, V. G., Lehmann, B. D. & Pietenpol, J. A. New strategies for triple-negative breast cancer-deciphering the heterogeneity. Clin. Cancer Res. 20, 782–790 (2014).PubMed PubMed Central CAS Article Google Scholar
- Gelmon, K. et al. Targeting triple-negative breast cancer: optimising therapeutic outcomes. Ann. Oncol. 23, 2223–2234 (2012).PubMed CAS Article Google Scholar
- Gagliato, D. de M., Jardim, D. L. F., Marchesi, M. S. P. & Hortobagyi, G. N. Mechanisms of resistance and sensitivity to anti-HER2 therapies in HER2+ breast cancer. Oncotarget 7, 64431–64446 (2016).Article Google Scholar
- Shah, S. P. et al. The clonal and mutational evolution spectrum of primary triple-negative breast cancers. Nature 486, 395–399 (2012).PubMed CAS Article Google Scholar
- Costa, R. et al. Targeting Epidermal Growth Factor Receptor in triple negative breast cancer: new discoveries and practical insights for drug development. Cancer Treat. Rev. 53, 111–119 (2017).PubMed CAS Article Google Scholar
- Swain, S. M. et al. Pertuzumab, trastuzumab, and docetaxel in HER2-positive metastatic breast cancer. N. Engl. J. Med. 372, 724–734 (2015).PubMed PubMed Central CAS Article Google Scholar
- Wright, M. H. et al. Brca1 breast tumors contain distinct CD44+/CD24- and CD133+cells with cancer stem cell characteristics. Breast Cancer Res. 10, R10 (2008).PubMed PubMed Central Article CAS Google Scholar
- Roberts, P. J. et al. Combined PI3K/mTOR and MEK inhibition provides broad antitumor activity in faithful murine cancer models. Clin. Cancer Res. 18, 5290–5303 (2012).PubMed PubMed Central CAS Article Google Scholar
- Hall, K., Lee, T.-H. & Aguilar, M.-I. The role of electrostatic interactions in the membrane binding of melittin. J. Mol. Recognit. 24, 108–118 (2011).PubMed CAS Article Google Scholar
- Rai, D. K., Qian, S. & Heller, W. T. The interaction of melittin with dimyristoyl phosphatidylcholine-dimyristoyl phosphatidylserine lipid bilayer membranes. Biochim. Biophys. Acta 1858, 2788–2794 (2016).PubMed CAS Article Google Scholar
- Krauson, A. J. et al. Conformational fine-tuning of pore-forming peptide potency and selectivity. J. Am. Chem. Soc. 137, 16144–16152 (2015).PubMed PubMed Central CAS Article Google Scholar
- Sharma, S. V. Melittin resistance—a counter selection for ras transformation. Oncogene 7, 193–201 (1992).PubMed CAS Google Scholar
- Morris, M. C., Deshayes, S., Heitz, F. & Divita, G. Cell-penetrating peptides: from molecular mechanisms to therapeutics. Biol. Cell 100, 201–217 (2008).PubMed CAS Article Google Scholar
- Dong, X., Hudson, N. E., Lu, C. & Springer, T. A. Structural determinants of integrin β-subunit specificity for latent TGF-β. Nat. Struct. Mol. Biol. 21, 1091–1096 (2014).PubMed PubMed Central CAS Article Google Scholar
- Li, W., Liu, C., Zhao, C., Zhai, L. & Lv, S. Downregulation of β3 integrin by miR-30a-5p modulates cell adhesion and invasion by interrupting Erk/Ets-1 network in triple-negative breast cancer. Int. J. Oncol. 48, 1155–1164 (2016).PubMed CAS Article Google Scholar
- Sorolla, A. et al. Triple-hit therapeutic approach for triple negative breast cancers using docetaxel nanoparticles, EN1-iPeps and RGD peptides. Nanomed. Nanotechnol. Biol. Med. 20, 102003 (2019).CAS Article Google Scholar
- Kretzmann, J. A. et al. Tumour suppression by targeted intravenous non-viral CRISPRa using dendritic polymers. Chem. Sci. 10, 7718–7727 (2019).PubMed PubMed Central CAS Article Google Scholar
- Kokot, G., Mally, M. & Svetina, S. The dynamics of melittin-induced membrane permeability. Eur. Biophys. J. 41, 461–474 (2012).PubMed CAS Article Google Scholar
- Carracedo, A. et al. Inhibition of mTORC1 leads to MAPK pathway activation through a PI3K-dependent feedback loop in human cancer. J. Clin. Investig. 118, 3065–3074 (2008).PubMed CAS PubMed Central Google Scholar
- Lee, T.-H. et al. Vascular endothelial growth factor mediates intracrine survival in human breast carcinoma cells through internally expressed VEGFR1/FLT1. PLoS Med. 4, e186 (2007).PubMed PubMed Central Article CAS Google Scholar
- Weigel, M. T. et al. Combination of imatinib and vinorelbine enhances cell growth inhibition in breast cancer cells via PDGFR β signalling. Cancer Lett. 273, 70–79 (2009).PubMed CAS Article Google Scholar
- Hall, M. P. et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 7, 1848–1857 (2012).PubMed PubMed Central CAS Article Google Scholar
- Machleidt, T. et al. NanoBRET—a novel BRET platform for the analysis of protein-protein interactions. ACS Chem. Biol. 10, 1797–1804 (2015).PubMed CAS Article Google Scholar
- Pfleger, K. D. G. & Eidne, K. A. Illuminating insights into protein-protein interactions using bioluminescence resonance energy transfer (BRET). Nat. Methods 3, 165–174 (2006).PubMed CAS Article Google Scholar
- Beltran, A. S., Graves, L. M. & Blancafort, P. Novel role of Engrailed 1 as a prosurvival transcription factor in basal-like breast cancer and engineering of interference peptides block its oncogenic function. Oncogene 33, 4767–4777 (2014).PubMed CAS Article Google Scholar
- Chou, T. C. & Talalay, P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 22, 27–55 (1984).CAS Article Google Scholar
- Prat, A. et al. Phenotypic and molecular characterization of the claudin-low intrinsic subtype of breast cancer. Breast Cancer Res. 12, R68 (2010).PubMed PubMed Central Article CAS Google Scholar
- Sorolla, A. et al. Sensitizing basal-like breast cancer to chemotherapy using nanoparticles conjugated with interference peptide. Nanoscale 8, 9343–9353 (2016).PubMed CAS Article Google Scholar
- Swoboda, A. & Nanda, R. Immune checkpoint blockade for breast cancer. Cancer Treat. Rev. 173, 155–165 (2018).Article Google Scholar
- Sharma, S. V. Melittin resistance: a counterselection for ras transformation. Oncogene 7, 193–201 (1992).PubMed CAS Google Scholar
- Barrajón-Catalán, E. et al. Selective death of human breast cancer cells by lytic immunoliposomes: correlation with their HER2 expression level. Cancer Lett. 290, 192–203 (2010).PubMed Article CAS Google Scholar
- Barnabas, N. & Cohen, D. Phenotypic and molecular characterization of MCF10DCIS and SUM breast cancer cell lines. Int. J. Breast Cancer 2013, 1–16 (2013).Article Google Scholar
- Hollestelle, A. et al. Distinct gene mutation profiles among luminal-type and basal-type breast cancer cell lines. Breast Cancer Res. Treat. 121, 53–64 (2010).PubMed Article Google Scholar
- Mota, A. et al. Molecular characterization of breast cancer cell lines by clinical immunohistochemical markers. Oncol. Lett. 13, 4708–4712 (2017).PubMed PubMed Central CAS Google Scholar
- Wu, G. J. et al. 17q23 amplifications in breast cancer involve the PAT1, RAD51C, PS6K, and SIGMA1B genes. Cancer Res. 60, 5371–5375 (2000).PubMed CAS Google Scholar
- Saal, L. H. et al. Recurrent gross mutations of the PTEN tumor suppressor gene in breast cancers with deficient DSB repair. Nat. Genet. 40, 102–107 (2008).PubMed CAS Article Google Scholar
- Liang, S. I. et al. Phosphorylated EGFR dimers are not sufficient to activate Ras. Cell Rep. 22, 2593–2600 (2018).PubMed PubMed Central CAS Article Google Scholar
- Lee, C., Bae, S.-J. S., Joo, H. & Bae, H. Melittin suppresses tumor progression by regulating tumor-associated macrophages in a Lewis lung carcinoma mouse model. Oncotarget 8, 54951–54965 (2017).PubMed PubMed Central Article Google Scholar
- Muenst, S. et al. Expression of programmed death ligand 1 (PD-L1) is associated with poor prognosis in human breast cancer. Breast Cancer Res. Treat. 146, 15–24 (2014).PubMed PubMed Central CAS Article Google Scholar
- Qin, T. et al. High PD-L1 expression was associated with poor prognosis in 870 Chinese patients with breast cancer. Oncotarget 6, 33972–33981 (2015).PubMed PubMed Central Article Google Scholar
- Zawlik, I. et al. Immune checkpoints in aggressive breast cancer subtypes. Neoplasma 63, 768–773 (2016).PubMed CAS Article Google Scholar
- Hou, Y., Nitta, H., Parwani, A. V. & Li, Z. PD-L1 and CD8 are associated with deficient mismatch repair status in triple-negative and HER2-positive breast cancers. Hum. Pathol. 86, 108–114 (2019).PubMed CAS Article Google Scholar
- Monypenny, J. et al. ALIX regulates tumor-mediated immunosuppression by controlling EGFR activity and PD-L1 presentation. Cell Rep. 24, 630–641 (2018).PubMed PubMed Central CAS Article Google Scholar
- Padmanabhan, R., Kheraldine, H. S., Meskin, N., Vranic, S. & Al Moustafa, A.-E. Crosstalk between HER2 and PD-1/PD-L1 in breast cancer: from clinical applications to mathematical models. Cancers 12, 636 (2020).PubMed Central CAS Article Google Scholar
- Polk, A., Svane, I.-M., Andersson, M. & Nielsen, D. Checkpoint inhibitors in breast cancer—current status. Cancer Treat. Rev. 63, 122–134 (2018).PubMed CAS Article Google Scholar
- Chaganty, B. K. R. et al. Trastuzumab upregulates PD-L1 as a potential mechanism of trastuzumab resistance through engagement of immune effector cells and stimulation of IFNγ secretion. Cancer Lett. 430, 47–56 (2018).PubMed PubMed Central CAS Article Google Scholar
- Cheng, B., Thapa, B., K. C., R. & Xu, P. Dual secured nano-melittin for the safe and effective eradication of cancer cells. J. Mater. Chem. B 3, 25–29 (2015).CAS Article Google Scholar
- Soman, N. R. et al. Molecularly targeted nanocarriers deliver the cytolytic peptide melittin specifically to tumor cells in mice, reducing tumor growth. J. Clin. Investig. 119, 2830–2842 (2009).PubMed CAS Article PubMed Central Google Scholar
- Sun, D. et al. The anti-cancer potency and mechanism of a novel tumor-activated fused toxin, DLM. Toxins 7, 423–438 (2015).PubMed PubMed Central CAS Article Google Scholar
- Sigismund, S., Avanzato, D. & Lanzetti, L. Emerging functions of the EGFR in cancer. Mol. Oncol. 12, 3–20 (2018).PubMed Article Google Scholar
- Iqbal, N. & Iqbal, N. Human epidermal growth factor receptor 2 (HER2) in cancers: overexpression and therapeutic implications. Mol. Biol. Int. 2014, 1–9 (2014).Article CAS Google Scholar
- Goding, J. Monoclonal Antibodies: Principles and Practice. (Academic Press, 1996).
- Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).PubMed CAS Article Google Scholar
- Stoddart, L. A. et al. Application of BRET to monitor ligand binding to GPCRs. Nat. Methods 12, 661–663 (2015).PubMed PubMed Central CAS Article Google Scholar
- Kilpatrick, L. E. et al. Real-time analysis of the binding of fluorescent VEGF 165 a to VEGFR2 in living cells: Effect of receptor tyrosine kinase inhibitors and fate of internalized agonist-receptor complexes. Biochem. Pharmacol. 136, 62–75 (2017).PubMed PubMed Central CAS Article Google Scholar
Poďakovanie
Táto práca bola podporená nasledujúcimi grantmi udelenými PB: Austrálska výskumná rada (ARC) Future Fellowship FT130101767, výskumné štipendium Cancer Council of Western Australia (CCWA) a projektové granty CCWA 1083745 a 1147435 a National Health and Medical Research Council ( NHMRC) udeľuje 1069308, 1147528 a 1165208. CD bolo podporené štipendiom austrálskeho vládneho výskumného vzdelávacieho programu (RTP) a štipendiom CCWA PhD Top Up Scholarship. AS oceňuje postdoktorandské štipendium od National Breast Cancer Foundation (PF-15-001) a Raine Foundation Priming Grant (RPG-004-19). E.J. bola podporená štipendiom ARC Industrial Transformation Training Center Fellowship (IC170100016). K.P. bol podporovaný NHMRC RD Wright Fellowship (1085842). Práca BRET bola čiastočne podporená grantom ARC LP160100857. Túto prácu podporili aj tieto granty udelené BB: granty ARC (LP100100438, DP130100087 a LP130100029), štipendium ARC Future Fellowship FT110100105 a grant FT110100528 a University of California Riverside. Autori by chceli oceniť podporu Jane Stout (Trinity College Dublin), Marka Browna (Royal Holloway, University of London), Kevina Li (FACS Facility na Harry Perkins Institute of Medical Research, UWA) a Paula Rigbyho (CMCA ).
Informácie o autorovi
Autori a príslušnosti
- School of Human Sciences, The University of Western Australia, Perth, WA, 6009, AustraliaCiara Duffy & Pilar Blancafort
- Cancer Epigenetics Group, Harry Perkins Institute of Medical Research, Perth, WA, 6009, AustraliaCiara Duffy, Anabel Sorolla, Edina Wang, Emily Golden, Eleanor Woodward & Pilar Blancafort
- Plant Energy Biology, The University of Western Australia, Perth, WA, 6009, AustraliaCiara Duffy
- Centre for Medical Research, The University of Western Australia, Perth, WA, 6009, AustraliaCiara Duffy, Anabel Sorolla, Edina Wang, Emily Golden, Eleanor Woodward, Kathleen Davern, Elizabeth Johnstone, Kevin Pfleger & Pilar Blancafort
- Monoclonal Antibody (MAb) Facility, Harry Perkins Institute of Medical Research, Perth, WA, 6009, AustraliaKathleen Davern & K. Swaminathan Iyer
- School of Molecular Sciences, The University of Western Australia, Perth, WA, 6009, AustraliaDiwei Ho
- Molecular Endocrinology and Pharmacology, Harry Perkins Institute of Medical Research, Perth, WA, 6009, AustraliaElizabeth Johnstone & Kevin Pfleger
- Australian Research Council Centre for Personalised Therapeutics Technologies, Perth, AustraliaElizabeth Johnstone & Kevin Pfleger
- Dimerix Limited; Nedlands, Perth, WA, 6009, AustraliaKevin Pfleger
- School of Medicine, The University of Western Australia, Perth, WA, 6009, AustraliaAndrew Redfern
- Centre for Integrative Bee Research (CIBER), Department of Entomology; University of California Riverside, Riverside, CA, 92521, USABoris Baer
- The Greehey Children’s Cancer Research Institute, The University of Texas Health Science Center at San Antonio, San Antonio, TX, 78229, USAPilar Blancafort
Príspevky
C.D. podieľal sa na návrhu a realizácii experimentov, analýze údajov, konštrukcii figúrok a napísal a redigoval článok. A.S. a E.W. prispeli k experimentom in vivo. napr. prispel k modelovaniu peptidov a ilustrácii plazmatickej membrány. E.W. prispel k imunofluorescenčným experimentom. K.D. prispeli k produkcii protilátok proti melitínu. D.H. prispel k rastrovacej elektrónovej mikroskopii. E.J. a K.P. prispel k vytvoreniu testov BRET. A.R. prezrel a upravil príspevok. K.S.I. prispel k dozoru a recenzoval príspevok. B.B. a P.B. prispel ku konceptualizácii, dohľadu a dizajnu experimentov a redigoval príspevok.
zodpovedajúci Autor
Korešpondencia s Pilar Blancafortovou.
Etické vyhlásenia
Konkurenčné záujmy
K.P. získala finančné prostriedky od spoločností Promega, BMG Labtech a Dimerix ako účastníckych organizácií ARC Linkage Grant. Tieto participujúce organizácie nezohrali žiadnu úlohu v žiadnom aspekte koncepcie alebo dizajnu výskumu, zberu, analýzy a interpretácie výsledkov, ani pri písaní a úprave príspevku. K.P. je hlavným vedeckým poradcom spoločnosti Dimerix, v ktorej vlastní podiel. Zostávajúci autori nedeklarujú žiadne konkurenčné záujmy.
Ďalšie informácie
Poznámka vydavateľa Springer Nature zostáva neutrálny, pokiaľ ide o jurisdikčné nároky vo zverejnených mapách a inštitucionálnych pridruženiach.
Práva a povolenia
Otvorený prístup Tento článok je licencovaný na základe medzinárodnej licencie Creative Commons Attribution 4.0, ktorá povoľuje používanie, zdieľanie, prispôsobovanie, distribúciu a reprodukciu na akomkoľvek médiu alebo formáte, ak uvediete pôvodného autora (autorov) a zdroj, uveďte odkaz na licenciu Creative Commons a uveďte, či boli vykonané zmeny. Obrázky alebo iné materiály tretích strán v tomto článku sú zahrnuté v licencii Creative Commons k článku, pokiaľ nie je v úverovom limite k materiálu uvedené inak. Ak materiál nie je zahrnutý v licencii Creative Commons k článku a vaše zamýšľané použitie nie je povolené zákonnými predpismi alebo prekračuje povolené použitie, budete musieť získať povolenie priamo od držiteľa autorských práv. Ak chcete zobraziť kópiu tejto licencie, navštívte stránku http://creativecommons.org/licenses/by/4.0/.
ZDROJ: https://www.nature.com/articles/s41698-020-00129-0#MOESM2