
Apiterapia je alternatívna terapia, ktorá sa opiera o využitie produktov včely medonosnej, predovšetkým včelieho jedu na liečbu mnohých ľudských chorôb. Jed sa môže dostať do ľudského tela manuálnou injekciou alebo priamym včelím bodnutím. Včelí jed obsahuje niekoľko aktívnych molekúl, ako sú peptidy a enzýmy, ktoré majú výhodný potenciál pri liečbe zápalov a ochorení centrálneho nervového systému, ako je Parkinsonova choroba, Alzheimerova choroba a amyotrofická laterálna skleróza. Okrem toho včelí jed preukázal sľubné výhody proti rôznym typom rakoviny, ako aj antivírusovú aktivitu, dokonca aj proti náročnému vírusu ľudskej imunodeficiencie (HIV). Mnohé štúdie opísali biologické aktivity zložiek včelieho jedu a spustili predklinické štúdie na zlepšenie potenciálneho využitia apitoxínu a jeho zložiek ako ďalšej generácie liekov. Cieľom tohto prehľadu je zhrnúť hlavné zlúčeniny včelieho jedu, ich primárne biologické vlastnosti, mechanizmy účinku a ich terapeutické hodnoty v alternatívnych terapeutických stratégiách.
Medzi včelami medonosnými je Apis mellifera (obrázok 1) hlavným druhom používaným na opeľovanie plodín na svete [1]. Používanie všetkých včelích produktov, vrátane včelieho jedu a medu, sa datuje tisíce rokov späť, keďže ich liečivé vlastnosti boli citované v náboženských knihách, ako je Biblia a Korán [2,3,4]. Apiterapia je odvetvie alternatívnej medicíny, ktoré sa opiera o používanie včelích produktov, ktoré pozostávajú z medu, peľu, propolisu, materskej kašičky a hlavne včelieho jedu (BV), ktorý je známy aj ako apitoxín [5,6].

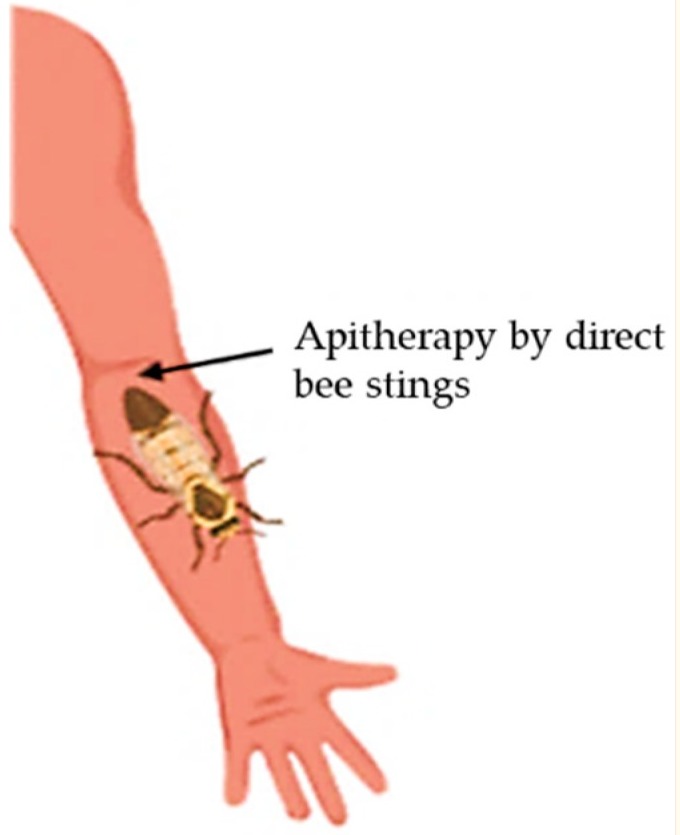
Terapia včelím jedom (BVT) je medicínska aplikácia BV z včiel medonosných do ľudského tela na liečbu niektorých chorôb, ako je reumatizmus artritída [7]. Táto stratégia sa používa v alternatívnej medicíne už viac ako 5000 rokov. Pozostáva buď z nepriamej aplikácie, extrakciou BV elektrickým stimulom s následnou injekciou do tela alebo priamo cez včelie bodnutie [8] (obrázok 2). Myšlienka využitia BV v medicíne vznikla z presvedčenia, že včelári takmer netrpia reumatizmom alebo problémami s kĺbmi.
BV je produkovaný samicami včiel a je známe, že obsahuje mnoho aktívnych zložiek vrátane: (i) peptidov, ako je melittín, apamín, peptid degranulujúci žírne bunky (MCD) a adolapín, (ii) enzýmy, ako je fosfolipáza A2 (PLA2) a hyaluronidáza a (iii) aminokyseliny a prchavé zlúčeniny. Niekoľko štúdií hodnotilo terapeutický potenciál týchto zložiek pri liečbe zápalových ochorení u ľudí, ako aj ochorení centrálneho nervového systému, ako je Parkinsonova choroba (PD), Alzheimerova choroba (AD) a amyotrofická laterálna skleróza (ALS), ako aj mnohé ďalšie stavy. [9,10]. Zaujímavé je, že včelí jed, podobne ako iné zvieracie jedy, tiež preukázal priaznivý protirakovinový a antivírusový potenciál proti rakovine vaječníkov a prostaty, ako aj HIV [11,12,13,14].
Včelí jed sa vyznačuje tým, že po uštipnutí vyvoláva alergické reakcie. Tieto reakcie sa môžu vyskytnúť v koži, dýchacom systéme, kardiovaskulárnom systéme a gastrointestinálnom systéme. Následne by ťažký anafylaktický šok mohol viesť k cerebrálnej alebo myokardiálnej ischémii [15,16]. Tieto alergické reakcie sú spôsobené prítomnosťou viacerých proteínových alergénov v jede, z ktorých väčšina má enzymatickú aktivitu [9]. Hlavné BV alergény a špecifické induktory imunoglobulínu E (IgE) sú PLA2, melitín a hyaluronidáza. Okrem mechanizmov sprostredkovaných IgE štúdie naznačujú, že alergény môžu zahŕňať aj reakcie nezávislé od IgE, ako je mediátor bradykinínu (BK), čo vedie k rôznym anafylaktickým symptómom [17,18]. Produkcia tohto neimunitného mediátora môže byť indukovaná melitínom, známym ako aktivátor PLA2, ktorý môže napodobňovať účinky BK na tracheálny tonus [17,19]. Okrem toho je MCD-peptid alebo peptid 401 schopný vyvolať anafylaktoidnú reakciu degranuláciou mastocytov [9,20].
Okrem molekulárnych štúdií, ktoré skúmajú možné mechanizmy za reakciou zápalových bodnutí včelou, mnohé klinické štúdie hlboko skúmajú potenciálne využitie BV na liečbu chronických ochorení. Preto je cieľom nasledujúcich častí prehľadu zdôrazniť primárne biologické vlastnosti BV a jeho bioaktívnych molekúl, ktoré majú potenciál pri vývoji terapeutických stratégií.
Hlavné zlúčeniny včelieho jedu
BV je priehľadná kvapalina bez zápachu obsahujúca hydrolytickú zmes bielkovín s kyslým pH (4,5 až 5,5), ktorú včely často používajú ako obranný nástroj proti predátorom. Jedna kvapka BV pozostáva z 88 % vody a len 0,1 µg suchého jedu [10]. Ten je mimoriadne komplexnou zmesou peptidov vrátane melittínu, adolapínu, apamínu a MCD-peptidu. Obsahuje tiež enzýmy, predovšetkým PLA2, a zlúčeniny s nízkou molekulovou hmotnosťou, ako sú bioaktívne amíny (napr. histamín a epinefrín) a minerály [9].
2.1. Melittin
Melittín, 26-zvyškový peptid, je hlavnou zložkou BV a tvorí 40–60 % jeho zloženia [21]. Karboxylová koncová oblasť peptidu je hydrofilná a zodpovedná za lytický účinok, zatiaľ čo aminokoncová oblasť jeho sekvencie je prevažne hydrofóbna bez lytickej aktivity [22]. Amfipatická vlastnosť melittínu ho robí rozpustným vo vode v monomérnej aj tetramérnej forme. Umožňuje tiež ľahké vloženie melitínu do membrán narušením prirodzených aj syntetických fosfolipidových dvojvrstiev. Predchádzajúce štúdie ukázali, že mechanizmus účinku melittínu pri rozrušovaní membrán je sprostredkovaný tvorbou pórov lýzujúcich prokaryotické aj eukaryotické bunky v neselektívnej hmote. V skutočnosti sa melitín viaže na membrány ako monoméry, ale pôsobí na membránu inkluzívne. V závislosti od koncentrácie môže tento biopeptid indukovať prechodné alebo stabilné póry [23]. Keď sa vytvorí prechodný pór, cez membránu sú schopné difundovať iba ióny. V prípade stabilnej tvorby pórov sa však membrána stáva priepustnou pre relatívne veľké molekuly, ako je glukóza [24]. Tvorba pórov vyvolaná melitínom je zodpovedná za jeho hemolytické, antimikrobiálne, antifungálne a protinádorové aktivity [12,25]. Nedávno sa ukázalo, že melitín spôsobuje nervové plastické zmeny pozdĺž dráh signalizácie bolesti aktiváciou a senzibilizáciou nociceptorových buniek. Mechanizmus zahŕňa fosforyláciu mitogénom aktivovaných proteínkináz (MAPK), ako aj aktiváciu tepelných nociceptívnych kanálov, ako je TRPV1 (prechodný receptorový potenciál vaniloidného receptora 1), ATP-gated P2X a P2Y purinergné receptory. Podobne môže melitín pôsobiť ako aktivátor PLA2 [26]. Je to tiež hlavná biologicky aktívna látka BV, ktorá po podaní do akupunktúry pacienta vyvoláva antinociceptívne, protizápalové a antiartritické účinky [27].
2.2. Apamin
Apamín je 18-aminokyselinový peptid obsahujúci dva disulfidové mostíky. Je to najmenší neurotoxín v BV [28]. Tento polypeptid je schopný prejsť hematoencefalickou bariérou, a preto ovplyvňuje fungovanie centrálneho nervového systému rôznymi spôsobmi účinku. Spôsobuje napríklad neurotoxické účinky v mieche cicavcov, čo má za následok hyperaktivitu a záchvaty, ako sa to dokázalo u potkanov. Blokovaním vápnikom aktivovaných K+ kanálov je apamín tiež schopný ovplyvniť priepustnosť bunkovej membrány voči draselným iónom (K+). V hladkom svalstve ciev je toxín schopný inhibovať proliferáciu a migráciu buniek hladkého svalstva ciev prostredníctvom signálnych dráh Akt a Erk [29]. Toto zistenie poukazuje na potenciál apamínu v stratégiách liečby aterosklerózy. Ďalšia štúdia hodnotila dôsledky citlivosti K+ kanálov na apamín a ukázala, že neurotoxín môže inhibovať NO-indukovanú relaxáciu spontánnej kontraktilnej aktivity myometria u netehotných žien [30].
2.3. Peptid degranulujúci žírne bunky (MCD).
MCD peptid, tiež známy ako peptid 401, je BV polypeptid obsahujúci 22 aminokyselín s podobnou štruktúrou ako apamín, pretože oba obsahujú dve disulfidové väzby. Tvorí 2–3 % sušiny BV. Názov MCD odráža biologický účinok pri uvoľňovaní histamínu zo žírnych buniek. Je to epileptogénny neurotoxín, dôležitý inhibítor K+ kanálov a môže spôsobiť významné zníženie krvného tlaku potkanov [31]. Zdá sa, že niektoré biologické aktivity MCD majú odlišné mechanizmy a môžu predstavovať dobrú ilustráciu vzťahu medzi štruktúrou a funkciou. Štúdie opisujú MCD ako silné protizápalové činidlo a môže slúžiť ako potenciálny kandidát na štúdium sekrečných mechanizmov zápalových buniek, ako sú mastocyty, bazofily a leukocyty, čo vedie k návrhu zlúčenín s terapeutickými aplikáciami [32].
2.4. Adolapin
Adolapín je bázický polypeptid so 103 aminokyselinovými zvyškami. Zodpovedá 1 % suchej hmotnosti BV. Výskumníci ukázali, že adolapín má protizápalové, antinociceptívne a antipyretické účinky tým, že blokuje syntézu prostaglandínov a inhibuje aktivitu cyklooxygenázy [33]. Polypeptid môže inhibovať lipoxygenázu z ľudských krvných doštičiek a môže mať analgetický účinok podľa Jung et al. [34].
2.5. Fosfolipáza A2
PLA2, najsmrteľnejší enzým v BV, je jeden polypeptidový reťazec 128 aminokyselín obsahujúci štyri disulfidové mostíky. Fosfolipáza A2 z včelieho jedu (bvPLA2) patrí do skupiny III enzýmov sPLA2 a môže pôsobiť ako ligand pre špecifické receptory. BvPLA2 predstavuje 12–15 % sušiny BV a je extrémne zásaditý. BvPLA2 je hydrolytický enzým, schopný špecificky štiepiť sn-2 acylovú väzbu fosfolipidov na rozhraní voda/lipid [35]. Je zaujímavé, že jeho činnosť môže byť zlepšená melittínom. Ukázalo sa, že k tomu dochádza počas procesu lýzy erytrocytov, čo dokazuje prítomnosť synergického účinku medzi bvPLA2 a melitínom [36,37]. V skutočnosti sa preukázalo, že melitín pomáha pri vystavovaní membránových fosfolipidov katalytickému miestu enzýmov prostredníctvom otvárania melitínom indukovaných kanálov [38]. Okrem toho nové experimentálne údaje preukázali ochranné imunitné reakcie bvPLA2 proti širokému spektru chorôb, ako je astma, Alzheimerova choroba a Parkinsonova choroba [39,40,41]. BvPLA2 hrá neuroprotektívnu úlohu tým, že indukuje mikrogliálnu deaktiváciu a znižuje infiltráciu CD4+ T buniek v MPTP-indukovanom myšom modeli PD (MPTP: 1-metyl-4-fenyl-1,2,3,6-tetrahydropyridín) [42].
2.6. Hyaluronidáza
Hyaluronidáza predstavuje 1,5–2 % sušiny BV a je známe, že rozkladá kyselinu hyalurónovú v tkanivách, ako napríklad v synoviálnej burze pri reumatoidnej artritíde. BV hyaluronidáza umožňuje aktívnym zložkám BV účinne difundovať do tkaniva obete ovplyvnením jeho štrukturálnej integrity a zvýšením prietoku krvi v danej oblasti. Tieto dve akcie sa kombinujú, aby zintenzívnili široké rozšírenie jedu [43,44].
Bioaktivity a terapeutické aplikácie včelieho jedu a jeho hlavných zlúčenín
3.1. Protizápalový potenciál
Zápal je ochranný proces pre telo v reakcii na škodlivé podnety. Chronický zápal môže viesť k rozvoju niekoľkých chorôb, ako je reumatoidná artritída (RA), cukrovka, kardiovaskulárne choroby, obezita, astma, kožné choroby a choroby súvisiace s CNS, ako sú PD, AD a ALS [45].
Melittin, keď sa podáva vo vysokých dávkach, spôsobuje lokálnu bolesť, svrbenie a zápal. Avšak nízke dávky tejto BV zlúčeniny môžu vyvolať široké protizápalové účinky. Mnohé správy skúmali protizápalové mechanizmy melitínu pri rôznych ochoreniach, ako je RA a ALS [46,47]. V skutočnosti pôsobí inhibíciou zápalových cytokínov, ako je interleukín-6 (IL-6), IL-8, tumor nekrotizujúci faktor-a (TNF-a) a interferón-y (IFN-y). Okrem toho melitín znižuje signálne dráhy, ktoré aktivujú zápalové cytokíny, vrátane nukleárneho faktora-kappa B (NF-KB), proteínkinázy Akt a kináz regulovaných extracelulárnym signálom (ERK1/2) v ľudských keratinocytoch ošetrených lipopolysacharidom porfyromonas gingivalis (PgLPS). Tieto zistenia naznačujú, že blokovaním ich primárnych signálnych dráh melitín inhibuje zápalové cytokíny, čo vedie k zníženiu zápalu v koži, pečeni, kĺboch a neurónovom tkanive [48].
Čo sa týka kožných ochorení, nedávna štúdia Kim et al. ukázali, že BV znižuje atopickú dermatitídu, najčastejšie alergické chronické zápalové ochorenie kože [49]. V skutočnosti jed stimuluje produkciu CD55 spustením dráh ERK1/2, čo vedie k zmierneniu symptómov ochorenia [50]. Je zaujímavé, že predchádzajúca štúdia Shina a kol. opísali protizápalový potenciál bvPLA2 pri kožných ochoreniach tým, že ukázali, že enzým tlmí atopický zápal kože prostredníctvom interakcie s CD206 [51].
3.2. Aplikácia BV na liečbu neurodegeneratívnych chorôb
3.2.1. Parkinsonova choroba PD je degeneratívna pohybová porucha, ktorá vedie u pacientov k progresívnej invalidite. Patologickými znakmi ochorenia sú progresívna strata dopaminergných neurónov v substantia nigra (štruktúra bazálnych ganglií nachádzajúca sa v ľudskom mozgu) a prítomnosť Lewyho teliesok, ktoré obsahujú agregáty alfa-synukleínu, široko distribuovaného proteínu v mozgu. [52,53]. Abnormálna mikrogliálna aktivácia je tiež patologickým znakom pri rôznych neurodegeneratívnych ochoreniach vrátane PD [54]. Mnoho predklinických štúdií skúmalo účinok BV na migráciu leukocytov alebo mikrogliálnu aktiváciu na zvieracích a bunkových modeloch. Ďalšie testy hodnotili neuroprotektívny potenciál BV akupunktúrnej terapie (BVA) proti rotenónom indukovanému oxidačnému stresu, neurozápalu a apoptóze na modeloch myší s PD [55]. Rotenón je pesticíd, ktorý môže ovplyvniť patofyziologické mechanizmy, ktoré sa podieľajú na PD [55]. Je zaujímavé, že BV preukázal svoju schopnosť zabrániť deplécii dopamínu po podaní rotenónu. Okrem toho sa lokomotorická aktivita znovu obnovila po ošetrení modelov myší PD pomocou BV. Liečba účinne potlačila poškodenie DNA a inhibovala expresiu apoptotických génov Bax, Bcl-2 a kaspázy-3 v mozgu PD myší. Tieto zistenia ukazujú, že BV normalizovala všetky apoptotické a neurozápalové markery a obnovila neurochémiu mozgu po poranení rotenónom [56]. Tiež sa ukázalo, že BV môže chrániť doparminergné neuróny pred degeneráciou v experimentálnych modeloch PD. Spolu s týmto zistením sa zistilo, že akupunktúrna stimulácia dolných zadných končatín BV má ochranný účinok na MPTP (1-metyl-4-fenyl-1,2,3,6-tetrahydropyridín) myšom modeli PD [57].
3.2.2. Alzheimerova choroba AD je najčastejším neurodegeneratívnym ochorením a na jeho vzniku sa podieľa mnoho patologických procesov [58]. Hypotéza amyloidnej kaskády a toxicita amyloidných beta (Aβ) peptidov však doteraz dominovala výskumu vďaka pokročilým štúdiám, ktoré ukazujú, že agregáty tohto peptidu sú charakteristické znaky ochorenia [59,60,61]. Hoci etiológia AD zostáva neznáma, dôkazy naznačujú, že zápalové odpovede môžu hrať kľúčovú úlohu v jej patogenéze [62,63]. Súčasné liečby kognitívnej straty súvisiacej s AD sa spoliehajú na použitie muskarínových alebo nikotínových receptorových ligandov a inhibítora acetylcholínesterázy (AChE) [64]. Ako alternatívnu stratégiu Ye et al. ukázali, že bvPLA2 možno použiť ako liečbu na blokovanie progresie AD u transgénnych myší [40]. Je to kvôli schopnosti bvPLA2 znižovať akumuláciu Ap a zlepšovať kognitívne funkcie v mozgoch myší. Rovnaká štúdia podobne ukazuje, že bvPLA2 môže zvýšiť metabolizmus glukózy v mozgu a znížiť neurozápalové reakcie v hipokampe, čo môže obmedziť patogenézu AD [40]. Nedávna štúdia tiež ukázala, že populácie regulačných T-buniek by mohli byť modulované liečbou bvPLA2 v myšom modeli 3xTg-AD. Preto autori navrhli nový terapeutický prístup na zníženie progresie AD kombináciou liečby bvPLA2 spolu s vakcinačnou terapiou Aβ, aby sa zabránilo jej nežiaducej zápalovej odpovedi [60].
3.2.3. Amyotrofická laterálna skleróza ALS je ochorenie CNS, ktoré spôsobuje smrť motorických neurónov [65]. Významnou črtou ALS je abnormálna akumulácia mutantných proteínových agregátov SOD1 (mtSOD1) [66]. Myší model ALS nesúci mutovaný gén mtSOD1 so substitúciou glycínu za alanín (SOD1G93A) charakterizoval Jaarsma et al., čím sa uľahčilo pochopenie etiológie ALS [67]. Štúdie in vitro aj in vivo s použitím mutantných transgénnych myší SOD1 preukázali rôzne bunkové patogénne udalosti v motorických neurónoch, ako je nesprávne poskladanie proteínov, dysfunkcia mitochondrií a akumulácia neurofilamentov [67]. Je zaujímavé, že BV preukázal určitý potenciál na pôsobenie proti tejto chorobe. V skutočnosti podávanie BV v presnom a symptomatickom štádiu progresie ALS vedie k zvýšeniu motorickej aktivity u myší s mutantom SOD1G93A a k predĺženiu očakávanej dĺžky života v porovnaní s kontrolnými myšami rovnakého veku. Mohlo by to byť spôsobené zablokovaním aktivovanej mikroglie, ktorá sa zvyčajne nachádza v myšacích modeloch ALS [68]. Ďalšia štúdia preukázala, že akupunktúra včelieho jedu (BVA) na ST36 inhibuje neurozápal v mieche myší so symptomatickou ALS signifikantným znížením hladín zápalových proteínov ako TLR4, CD14 a TNF-α [69].
3.3. Aplikácie BV a/alebo Melittinu pri rakovine
Použitie apitoxínu, najmä jeho hlavnej zlúčeniny melittínu, ako novej stratégie liečby rakoviny nadobudlo v poslednej dobe veľký význam [70,71]. V skutočnosti je známe, že melitín je nešpecifický cytolytický peptid, ktorý môže atakovať lipidovú dvojvrstvu, čo vedie k významnej toxicite pri intravenóznej injekcii [72]. Napriek tomu sa využilo mnoho optimalizačných prístupov, vrátane použitia dodávania melitínu na báze nanočastíc. Je pozoruhodné, že surový BV, ako aj melitín preukázali protinádorové aktivity proti rôznym typom rakovinových buniek vrátane buniek rakoviny prsníka, pečene, leukémie, pľúc, melanómu a prostaty [70,72,73,74,75]. Wang a kol. [76] skúmali mechanizmus protinádorovej aktivity melitínu a ukázali, že melitín môže indukovať apoptózu buniek hepatocelulárneho karcinómu (HCC) prostredníctvom aktivácie signálnej dráhy CAMKII-TAK1-JNK/p38 (CAMKII: Ca2+/kalmodulín-dependentná proteínkináza; TAK1 : Transformujúci rastový faktor-beta-aktivovaná kináza 1; JNK/p38: Mitogénom-aktivované proteínkinázy). Okrem toho môže melitín senzibilizovať TRAIL-rezistentné bunky HCC (TRAIL: ligand indukujúci apoptózu súvisiaci s faktorom nekrotizujúcim nádory) na apoptózu indukovanú TRAIL, pravdepodobne prostredníctvom aktivácie dráhy CAMKII-TAK1-JNK/p38 a inhibície dráhy IKK-NFκB (obrázok 3). Tieto zistenia sú v súlade s aktiváciou vápnikových kanálov melitínom, ktorá vedie k zvýšeniu intracelulárnej koncentrácie Ca2+ a aktivácii CaMKII citlivého na vápnik, ako je vidieť aj na obrázku 3 [76].
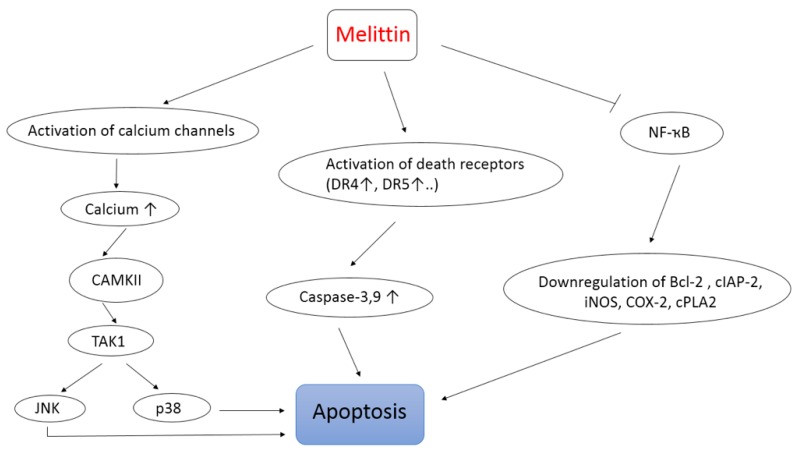
Park a kol. [13] tiež uviedli, že BV a jeho hlavná zložka, melitín, indukujú inhibíciu rastu rakovinových buniek in vitro aj in vivo prostredníctvom aktivácie kaspázových dráh (3 a 9) a inhibíciou signalizácie NF-κB a jej downstream proliferácie a antiapoptotické génové produkty ako Bcl-2, cIAP-2, iNOS, COX-2 a cPLA2 (obrázok 3) [13]. Podobne Zheng a kol. [77] preukázali, že BV má antiproliferatívny účinok a indukuje apoptózu prostredníctvom aktivácie receptorov smrti (DR4 a DR5). Ďalšie zaujímavé zistenie sa objavilo o melitíne zdôraznením jeho antimetastatických a protirastových vlastností [73]. Pri rakovine sú metastázy a invázia malígnych buniek hlavnými príčinami progresie ochorenia. Preto sa výskumníci v oblasti rakoviny zameriavajú na pochopenie molekulárnych mechanizmov, ktoré regulujú migráciu malígnych buniek a možný spôsob, ako tomu zabrániť, ako rozhodujúci krok v ich boji proti rakovine [78,79]. V tejto súvislosti sa zistilo, že melitín inhibuje in vitro a in vivo motilitu buniek HCC potlačením dráh závislých od Rac1 [73]. Na druhej strane nedávna štúdia dokázala, že kombinácia melittínu s chemoterapeutikom, ako je temozolomid, výrazne znižuje rast spolu s inváziou buniek melanómu v porovnaní so stavmi, keď sa TMZ alebo melitín používali samostatne [71].
Tieto zistenia poukazujú na veľký potenciál melittínu pri liečbe rakoviny pôsobením na rôzne kľúčové body choroby a mali by sa ďalej rozoberať.
Napriek presvedčivým údajom týkajúcim sa potenciálneho použitia BV, konkrétnejšie melittínu, proti rôznym typom rakoviny, jeho použiteľnosť u ľudí zostáva veľmi náročná z dôvodu jeho nešpecifickej cytotoxicity [80]. Súčasné optimalizačné metódy sa zameriavajú na dodávanie melittínu na báze nanočastíc, aby sa predišlo takýmto problémom. Vďaka nanotechnológii bolo možné vyvinúť a efektívne otestovať konjugáty melittínu proti širokému spektru ľudských typov rakoviny v predklinických modeloch [81]. Cheng a kol. zamerané na vývoj účinného a zároveň bezpečného systému dodávania melittínu, ktorý môže znížiť jeho hemolytickú aktivitu pri zachovaní jeho cytotoxických výhod. Preto bol navrhnutý duálny zabezpečený nano-žihadlo (DSNS) prostredníctvom kombinácie zwitteriónového glykolchitosanu a disulfidových väzieb. DSNS nabitý melittínom vykazoval takmer úplný cytotoxický účinok na mnohé typy rakovinových buniek pri veľmi nízkych koncentráciách, pričom červené krvinky zostali nepoškodené [82]. Okrem toho sa ukázalo, že intravenózne podanie nanočastíc s prekurzorom melittínu s použitím nanočastíc per fluorouhlík v modeli myší s melanómom účinne znížilo rýchlosť rastu nádoru v porovnaní s liečbou fyziologickým roztokom a slepými nanočasticami [83].
3.4. Antivírusové a antibakteriálne vlastnosti
Je dobre známe, že BV a jej dve hlavné zložky (melitín a PLA2) vykazujú antimikrobiálne aktivity, a preto sa môžu použiť ako doplnkové antibakteriálne látky [84,85,86,87]. Tieto zlúčeniny uplatňujú svoje účinky proti baktériám indukciou pórov cez ich membrány, čo vedie k ich štiepeniu a následne lýze [36].
Napriek tomu sa antivírusový účinok BV v literatúre príliš nespomína. Nedávna štúdia skúmala BV antivírusový potenciál a priniesla zaujímavé zistenia in vivo aj in vitro. Táto štúdia ukázala, že BV a melitín majú významné antivírusové účinky proti mnohým obaleným vírusom (vírus vezikulárnej stomatitídy, vírus chrípky A, vírus herpes simplex atď.) a neobaleným vírusom (enterovírus-71 a vírus coxsackie) in vitro [88]. Štúdia tiež ukázala, že melitín chránil myši, ktoré boli vystavené smrteľným dávkam vírusu chrípky A H1N1. Hoci presný mechanizmus účinku, ktorým BV a melitín pôsobia ako antivírusové činidlá, zostáva nejasný, potvrdilo sa, že BV interaguje priamo s vírusovým povrchom. Okrem toho môže BV a jeho zložky stimulovať interferón typu I (IFN), a teda potlačiť replikáciu vírusu v hostiteľskej bunke [89].
Okrem toho výskumníci z Washington University School of Medicine v St. Louis oznámili možnú aplikáciu nanočastíc naplnených melittínom pri ničení vírusu ľudskej imunodeficiencie, pričom neinfikované bunky zostávajú bez poškodenia. V tomto prístupe autori navrhujú preventívnu stratégiu, v ktorej sa tieto nanočastice používajú pri vývoji vaginálneho gélu, ktorý inhibuje šírenie HIV. Jeho teoretický princíp je nasledovný: Molekuly melittínu prítomné na nanočasticiach sa spájajú s vírusovým obalom a vytvárajú pórovité útočné komplexy, čím sa vírusový obal rozbíja [14]. Ďalšia štúdia ukázala, že bvPLA2 môže tiež blokovať replikáciu vírusu. Ten istý tím ďalej identifikoval peptidovú sekvenciu bvPLA2 zodpovednú za inhibíciu replikácie HIV [89,90,91,92].
- Závery
Použitie BV na lekárske účely možno vysledovať tisíce rokov späť. Tu sa diskutuje o terapeutickom záujme surového včelieho jedu a/alebo jeho hlavných zlúčenín, najmä melitínu. Ten poskytuje široké protizápalové vlastnosti tým, že ovplyvňuje signálne dráhy primárneho zápalu a indukuje inhibíciu expresie prozápalových génov. BV má tiež neuroprotektívny potenciál pri neurodegeneratívnych ochoreniach, ako sú PD, AD a ALS tým, že významne blokuje ich progresiu a zlepšuje kognitívne fungovanie na myšacích modeloch. Pokiaľ ide o protinádorovú aktivitu, melitín aj BV majú cytotoxický účinok na rakovinové bunky a významnú antimetastatickú aktivitu. Optimalizačné prístupy sa v súčasnosti zameriavajú na možné využitie nanočasticového dodávania melittínu alebo dokonca BV, aby sa predišlo ich nešpecifickému cytotoxickému účinku. Antivírusová aktivita BV je tiež sľubná, pretože BV a melitín majú pozoruhodné toxické účinky proti širokému spektru obalených vírusov, vrátane náročného HIV, a niekoľkých neobalených vírusov. Napokon, klinická aplikácia terapie BV je ešte dlhá cesta, ale výskumníci veria, že prebiehajúca práca na tejto téme nakoniec umožní, aby sa BV a jej zlúčeniny v nadchádzajúcich rokoch považovali za konečných kandidátov na rôzne terapie.
Poďakovanie
Autori by sa chceli poďakovať Eliemu N. Mahfoudovi za usilovnú korektúru príspevku. Za užitočnú diskusiu ďakujú aj Cesarovi Matteiovi a Christianovi Legrosovi.
Autorské príspevky
Konceptualizácia, Z.F. a J.-M.S.; získavanie financií, Z.F. a D.E.O.; písanie – príprava pôvodného návrhu, R.W.; písanie – recenzia a úprava, J.F., R.W., M.R. a Z.F.
Financovanie
Tento výskum financovala Libanonská univerzita.
Konflikt záujmov
Autori nedeklarujú žiadny konflikt záujmov.
Referencie
1. Greenleaf S.S., Kremen C. Wild bees enhance honey bees’ pollination of hybrid sunflower. Proc. Natl. Acad. Sci. USA. 2006;103:13890–13895. doi: 10.1073/pnas.0600929103. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
2. Fratellone P.M. Apitherapy Products for Medicinal Use. J. Nutr. Food Sci. 2015;5 doi: 10.1089/acm.2015.0346. [PubMed] [CrossRef] [Google Scholar]
3. Ali M.A.A.-S.M. Studies on bee venom and its medical uses. Int. J. Adv. Res. Technol. 2012;1:69–83. [Google Scholar]
4. El-Wahab S.D.A., Eita L.H. The effectiveness of live bee sting acupuncture on depression. J. Nurs. Health Sci. 2015;4:19–27. [Google Scholar]
5. Trumbeckaite S., Dauksiene J., Bernatoniene J., Janulis V. Knowledge, attitudes, and usage of apitherapy for disease prevention and treatment among undergraduate pharmacy students in Lithuania. Evid.-Based Complement. Altern. Med. 2015;9 doi: 10.1155/2015/172502. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. Hellner M., von Georgi D.W.R., Münstedt K. Apitherapy: Usage and experience in German beekeepers. Evid.-Based Complement. Altern. Med. 2007;5:475–479. doi: 10.1093/ecam/nem052. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
7. Lee J.A., Son M.J., Choi J., Yun K.J., Jun J.H., Lee M.S. Bee venom acupuncture for rheumatoid arthritis: A systematic review protocol. Bmj Open. 2014;4 doi: 10.1136/bmjopen-2013-004602. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
8. Zhang S., Liu Y., Ye Y., Wang X.R., Lin L.T., Xiao L.Y., Zhou P., Shi G.X., Liu C.Z. Bee venom therapy: Potential mechanisms and therapeutic applications. Toxicon. 2018;148:64–73. doi: 10.1016/j.toxicon.2018.04.012. [PubMed] [CrossRef] [Google Scholar]
9. Moreno M., Giralt E. Three Valuable Peptides from Bee and Wasp Venoms for Therapeutic and Biotechnological Use: Melittin, Apamin and Mastoparan. Toxins. 2015;7:1126–1150. doi: 10.3390/toxins7041126. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
10. Bellik Y. Bee Venom: Its potential use in alternative medicine. Anti-Infect. Agents. 2015;13:3–16. doi: 10.2174/2211352513666150318234624. [CrossRef] [Google Scholar]
11. Jo M., Park M.H., Kollipara P.S., An B.J., Song H.S., Han S.B., Kim J.H., Song M.J., Hong J.T. Anti-cancer effect of bee venom toxin and melittin in ovarian cancer cells through induction of death receptors and inhibition of JAK2/STAT3 pathway. Toxicol. Appl. Pharmacol. 2012;258:72–81. doi: 10.1016/j.taap.2011.10.009. [PubMed] [CrossRef] [Google Scholar]
12. Rady I., Siddiqui I.A., Rady M., Mukhtar H. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. Cancer Lett. 2017;402:16–31. doi: 10.1016/j.canlet.2017.05.010. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Park M.H., Choi M.S., Kwak D.H., Oh K.W., Yoon D.Y., Han S.B., Song H.S., Song M.J., Hong J.T. Anti-cancer effect of bee venom in prostate cancer cells through activation of caspase pathway via inactivation of NF-κB. Prostate. 2011;71:801–812. doi: 10.1002/pros.21296. [PubMed] [CrossRef] [Google Scholar]
14. Hood J.L., Jallouk A.P., Campbell N., Ratner L., Wickline S.A. Cytolytic nanoparticles attenuate HIV-1 infectivity. Antivir. Ther. 2013;18:95–103. doi: 10.3851/IMP2346. [PubMed] [CrossRef] [Google Scholar]
15. Bilò M.B., Bonifazi F. The natural history and epidemiology of insect venom allergy: Clinical implications. Clin. Exp. Allergy. 2009;39:1467–1476. doi: 10.1111/j.1365-2222.2009.03324.x. [PubMed] [CrossRef] [Google Scholar]
16. Golden D.B.K. Insect sting anaphylaxis. Immunol. Allergy Clin. North. Am. 2007;27:261. doi: 10.1016/j.iac.2007.03.008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
17. Mingomataj E.Ç., Bakiri A.H. Episodic hemorrhage during honeybee venom anaphylaxis: Potential mechanisms. J. Investig. Allergol. Clin. Immunol. 2012;22:237–244. [PubMed] [Google Scholar]
18. Kaplan A.P. Kinins, airway obstruction, and anaphylaxis. Chem. Immunol. Allergy. 2010;95:67–84. [PubMed] [Google Scholar]
19. Mingomataj E., Bakiri A. The inhibition of kallikrein-bradykinin pathway may be useful in the reduction of allergic reactions during honeybee venom immunotherapy. J. Med. Hypotheses Ideas. 2009;3:10. [Google Scholar]
20. Ziai M.R., Russek S., Wang H.C., Beer B., Blume A.J. Mast cell degranulating peptide: A multi-functional neurotoxin. J. Pharm. Pharmacol. 1990;42:457–461. doi: 10.1111/j.2042-7158.1990.tb06595.x. [PubMed] [CrossRef] [Google Scholar]
21. Chen J., Guan S.M., Sun W., Fu H. Melittin, the Major Pain-Producing Substance of Bee Venom. Neurosci Bull. 2016;32 doi: 10.1007/s12264-016-0024-y. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
22. Raghuraman H., Chattopadhyay A. Melittin: A membrane-active peptide with diverse functions. Biosci. Rep. 2007;27:189–223. doi: 10.1007/s10540-006-9030-z. [PubMed] [CrossRef] [Google Scholar]
23. Pino-Angeles A., Lazaridis T. Effects of peptide charge, orientation, and concentration on melittin transmembrane pores. Biophysj. J. 2018;114:2865–2874. doi: 10.1016/j.bpj.2018.05.006. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
24. Lee M.T., Sun T.L., Hung W.C., Huang H.W. Process of inducing pores in membranes by melittin. PNAS. 2013;110:14243–14248. doi: 10.1073/pnas.1307010110. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
25. Zarrinnahad H., Mahmoodzadeh A., Hamidi M.P., Mahdavi M., Moradi A., Bagheri K.P., Shahbazzadeh D. Apoptotic effect of melittin purified from iranian honey bee venom on human cervical cancer heLa cell line. Intj. Pept Res. Ther. 2018;24:563–570. doi: 10.1007/s10989-017-9641-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Keith D.J., Eshleman A.J., Janowsky A. Melittin stimulates fatty acid release through non-phospholipase-mediated mechanisms and interacts with the dopamine transporter and other membrane spanning proteins. Eur. J. Pharmacol. 2011;650:501–510. doi: 10.1016/j.ejphar.2010.10.023. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
27. Bae G., Bae H. Anti-Inflammatory applications of melittin, a major component of bee venom: Detailed mechanism of action and adverse effects. Molecules. 2016;21:616. [PMC free article] [PubMed] [Google Scholar]
28. Son D.J., Lee J.W., Lee Y.H., Song H.S., Lee C.K., Hong J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharm. Ther. 2007;115:246–270. doi: 10.1016/j.pharmthera.2007.04.004. [PubMed] [CrossRef] [Google Scholar]
29. Kim J.Y., Kim K.H., Lee W., An H.J., Lee S.J., Han S.M., Lee K.G., Park Y.Y., Kim K.S., Lee Y.S., et al. Apamin inhibits PDGF-BB-induced vascular smooth muscle cell proliferation and migration through suppressions of activated Akt and Erk signaling pathway. Vasc. Pharmacol. 2015;70 doi: 10.1016/j.vph.2014.12.004. [PubMed] [CrossRef] [Google Scholar]
30. Modzelewska B., Kostrzewska A., Sipowicz M., Kleszczewski T., Batra S. Apamin inhibits NO-induced relaxation of the spontaneous contractile activity of the myometrium from non-pregnant women. Reprod. Biol. Endocrinol. 2003;1:8. [PMC free article] [PubMed] [Google Scholar]
31. Hanson J.M., Morley J. Anti-inflammatory property of 401 (MCD-peptide), a peptide from the venom of the bee Apis mellifera (L.) Br. J. Pharmacol. 1974;50:383–392. doi: 10.1111/j.1476-5381.1974.tb09613.x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
32. Banks B.E., Dempsey C.E., Vernon C.A., Warner J.A., Ymey J. Anti-inflammatory activity of bee venom peptide 401 (mast cell degranulating peptide) and compound 48/80 results from mast cell degranulation in vivo. Br. J. Pharmacol. 1990;99:350–354. doi: 10.1111/j.1476-5381.1990.tb14707.x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Cherniack E.P., Govorushko S. To bee or not to bee: The potential efficacy and safety of bee venom acupuncture in humans. Toxicon. 2018;154:74–78. doi: 10.1016/j.toxicon.2018.09.013. [PubMed] [CrossRef] [Google Scholar]
34. Jung S.Y., Lee K.W., Choi S.M., Yang E.J. Bee Venom Protects against Rotenone-Induced Cell Death in NSC34 Motor Neuron Cells. Toxins. 2015;7:3715–3726. doi: 10.3390/toxins7093715. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
35. Samel M., Vija H., Kurvet I., Künnis-Beres K., Trummal K., Subbi J., Kahru A., Siigur J. Interactions of PLA2-s from Vipera lebetina, Vipera berus berus and Naja naja oxiana Venom with Platelets, Bacterial and Cancer Cells. Toxins. 2013;5:203–223. doi: 10.3390/toxins5020203. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
36. Leandro L.F., Mendes C.A., Casemiro L.A., Vinholis A.H., Cunha W.R., De Almeida R., Martins C.H. Antimicrobial activity of apitoxin, melittin and phospholipase A₂ of honey bee (Apis mellifera) venom against oral pathogens. An. Acad. Bras. Cienc. 2015;87:147–155. doi: 10.1590/0001-3765201520130511. [PubMed] [CrossRef] [Google Scholar]
37. Frangieh J., Salma Y., Haddad K., Mattei C., Legros C., Fajloun Z., El Obeid D. First Characterization of The Venom from Apis mellifera syriaca, A Honeybee from The Middle East Region. Toxins. 2019;11:191. doi: 10.3390/toxins11040191. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
38. Lu Z.M., Xie F., Fu H., Liu M.G., Cao F.L., Hao J., Chen J. Roles of peripheral P2X and P2Y receptors in the development of melittin-induced nociception and hypersensitivity. Neurochem. Res. 2008;33:2085–2091. doi: 10.1007/s11064-008-9689-6. [PubMed] [CrossRef] [Google Scholar]
39. Park S., Baek H., Jung K.H., Lee G., Lee H., Kang G.H., Lee G., Bae H. Bee venom phospholipase A2 suppresses allergic airway inflammation in an ovalbumin-induced asthma model through the induction of regulatory T cells. Immun. Inflamm. Dis. 2015;3:386–397. doi: 10.1002/iid3.76. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
40. Ye M., Chung H.S., Lee C., Yoon M.S., Yu A.R., Kim J.S., Hwang D.S., Shim I., Bae H. Neuroprotective effects of bee venomphospholipase A2 in the 3xTg AD mousemodel of Alzheimer’s disease. J. Neuroinflamm. 2016;13:10. [PMC free article] [PubMed] [Google Scholar]
41. Kim K.H., Lee S.Y., Shin J., Hwang J.T., Jeon H.N., Bae H. Dose-Dependent Neuroprotective Effect of Standardized Bee Venom Phospholipase A2 Against MPTP-Induced Parkinson’s Disease in Mice. Front. Aging Neurosci. 2019;11 doi: 10.3389/fnagi.2019.00080. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
42. Chung E.S., Lee G., Lee C., Ye M., Chung H.S., Kim H., Bae S.J., Hwang D.S., Bae H. Bee venom phospholipase A2, a novel Foxp3+ regulatory T cell inducer, protects dopaminergic neurons by modulating neuroinflammatory responses in a mouse model of Parkinson’s Disease. J. Immunol. 2015;195:4853–4860. doi: 10.4049/jimmunol.1500386. [PubMed] [CrossRef] [Google Scholar]
43. Topchiyeva T., Mammadova F.Z. The seasonal activity of hyaluronidase in venom of a honey bee (Apis mellifera L. caucasica) in various regions of Azerbaijan. J. Entomol. Zool. Stud. 2016;4:1388–1391. [Google Scholar]
44. Hossen M.S., Shapla U.M., Gan S.H., Khalil M.I. Impact of Bee Venom Enzymes on Diseases and Immune Responses. Molecules. 2017;22:25. doi: 10.3390/molecules22010025. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
45. Glass C.K., Saijo K., Winner B., Marchetto M.C., Gage F.H. Mechanisms Underlying Inflammation in Neurodegeneration. Cell. 2010;140:918–934. doi: 10.1016/j.cell.2010.02.016. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
46. Khalil W.S., Khalil E.A.G. Immune response modifying effects of bee venom protein [Melittin]/Autoclaved, L. donovani complex in CD1 Mice: The search for new vaccine adjuvants. J. Vaccines Vaccin. 2017;8:2. [Google Scholar]
47. Park H.J., Son D.J., Oh K.W., Kim K.H., Song H.S., Kim G.J., Oh G.T., Yoon D.Y., Hong J.T. Inhibition of inflammation mediator generation by suppression of NF-kB through interaction with the p50 subunit. Arthritis Rheum. 2004;50:504–3515. doi: 10.1002/art.20626. [PubMed] [CrossRef] [Google Scholar]
48. Kim W.H., An H.J., Kim J.Y., Gwon M.G., Gu H., Jeon M., Kim M.K., Han S.M., Park K.K. Anti-Inflammatory Effect of Melittin on Porphyromonas Gingivalis LPS-Stimulated Human Keratinocytes. Molecules. 2018;23:332. doi: 10.3390/molecules23020332. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
49. Ong P.Y., Leung D.Y.M. Bacterial and viral infections in atopic dermatitis: A comprehensive review. Clin. Rev. Allergy Immunol. 2016;51:329–337. doi: 10.1007/s12016-016-8548-5. [PubMed] [CrossRef] [Google Scholar]
50. Kim Y., Lee Y.W., Kim H., Chung D.K. Bee venom alleviates atopic dermatitis symptoms through the upregulation of decay-accelerating factor (DAF/CD55) Toxins. 2019;11:239. doi: 10.3390/toxins11050239. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
51. Shin D., Choi W., Bae H. Bee venom Phospholipase A2 alleviate house dust mite-induced atopic dermatitis-like skin lesions by the CD206 mannose receptor. Toxins. 2018;10:146. doi: 10.3390/toxins10040146. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
52. Goldman J.G., Williams-Gray C., Barker R.A., Duda J.E., Galvin J.E. The spectrum of cognitive impairment in Lewy body diseases. Mov. Disord. 2014;29:217–231. doi: 10.1002/mds.25866. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
53. Aarsland D., Creese B., Politis M., Chaudhuri K.R., Ffytche D.H., Weintraub D., Ballard C. Cognitive decline in Parkinson disease. Nat. Rev. Neurol. 2017;13:217–231. doi: 10.1038/nrneurol.2017.27. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
54. Iakovakis D., Hadjidimitriou S., Charisis V., Bostantzopoulou S., Katsarou Z., Hadjileontiadis L.J. Touchscreen typing-pattern analysis for detecting fine motor skills decline in early-stage Parkinson’s disease. Sci. Rep. 2018 doi: 10.1038/s41598-018-25999-0. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
55. Tanner C.M., Kamel F., Ross G.W., Hoppin J.A., Goldman S.M., Korell M., Marras C., Bhudhikanok G.S., Kasten M., Chade A.R., et al. Rotenone, paraquat, and Parkinson’s disease. Environ. Health Perspect. 2011;119:866–872. doi: 10.1289/ehp.1002839. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
56. Khalil W.K.B., Assaf N., ElShebiney S.A., Salem N.A. Neuroprotective effects of bee venom acupuncture therapy against rotenone-induced oxidative stress and apoptosis. Neurochem. Int. 2015;80:79–86. doi: 10.1016/j.neuint.2014.11.008. [PubMed] [CrossRef] [Google Scholar]
57. Alvarez-Fischer D., Noelker C., Vulinović F., Grünewald A., Chevarin C., Klein C., Oertel W.H., Hirsch E.C., Michel P.P., Hartmann A. Bee Venom and Its Component Apamin as Neuroprotective Agents in a Parkinson Disease Mouse Model. PLoS ONE. 2013:8. doi: 10.1371/journal.pone.0061700. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
58. Aksoz E., Gocmez S., Sahin T.D., Aksit D., Aksit H., Utkan T. The protective effect of metformin in scopolamine-induced learning and memory impairment in rats. Pharm. Rep. 2019;71:818–825. doi: 10.1016/j.pharep.2019.04.015. [PubMed] [CrossRef] [Google Scholar]
59. Sabri O., Sabbagh M.N., Seibyl J., Barthel H., Akatsu H., Ouchi Y., Senda K., Murayama S., Ishii K., Takao M., et al. Florbetaben PET imaging to detect amyloid beta plaques in Alzheimer’s disease: Phase 3 study. Alzheimers Dement. 2015;11:964–974. doi: 10.1016/j.jalz.2015.02.004. [PubMed] [CrossRef] [Google Scholar]
60. Baek H., Lee C., Choi D.B., Kim N.S., Kim Y.S., Ye Y.J., Kim Y.S., Kim J.S., Shim I., Bae H. Bee venom phospholipase A2 ameliorates Alzheimer’s disease pathology in Aβ vaccination treatment without inducing neuro-inflammation in a 3xTg-AD mouse model. Sci. Rep. 2018;8 doi: 10.1038/s41598-018-35030-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
61. Kakuda N., Miyasaka T., Iwasaki N., Nirasawa T., Wada-Kakuda S., Takahashi-Fujigasaki J., Murayama S., Ihara Y., Ikegawa M. Distinct deposition of amyloid-β species in brains with Alzheimer’s disease pathology visualized with MALDI imaging mass spectrometry. Acta Neuropathol. Commun. 2017;5 doi: 10.1186/s40478-017-0477-x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
62. Van Eldik L.J., Carrillo M.C., Cole P.E., Feuerbach D., Greenberg B., Hendrix J.A., Kennedy M., Kozauer N., Margolin R.A., Molinuevo J.L., et al. The roles of inflammation and immune mechanisms in Alzheimer’s disease. Alzheimers Dement. 2016;2:99–109. doi: 10.1016/j.trci.2016.05.001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
63. Kinney J.W., Bemiller S.M., Murtishaw A.S., Leisgang A.M., Salazar A.M., Lamb B.T. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimers Dement. 2018;4:575–590. doi: 10.1016/j.trci.2018.06.014. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
64. Terry A.V., Buccafusco J.J. The Cholinergic Hypothesis of Age and Alzheimer’s Disease-Related Cognitive Deficits: Recent Challenges and TheirImplications for Novel Drug Development. J. Pharm. Exp. Ther. 2003;306:821–827. doi: 10.1124/jpet.102.041616. [PubMed] [CrossRef] [Google Scholar]
65. Rajagopalan V., Pioro E.P. Unbiased MRI analyses identify micropathologic differences between upper motor neuron-predominant ALS phenotypes. Front. Neurosci. 2019;13 doi: 10.3389/fnins.2019.00704. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
66. Boillée S., Yamanaka K., Lobsiger C.S., Copeland N.G., Jenkins N.A., Kassiotis G., Kollias G., Cleveland D.W. Onset and progression in inherited ALS determined by motor neurons and microglia. Science. 2006;312:1389–1392. doi: 10.1126/science.1123511. [PubMed] [CrossRef] [Google Scholar]
67. Jaarsma D., Haasdijk E.D., Grashorn J.A.C., Hawkins R., Van Duijn W., Verspaget W.H., London J., Holstege J.C. Human Cu/Zn Superoxide Dismutase (SOD1) Overexpression in Mice Causes Mitochondrial Vacuolization, Axonal Degeneration, and Premature Motoneuron Death and Accelerates MotoneuronDisease in Mice Expressing a Familial AmyotrophicLateral Sclerosis Mutant SOD1. Neurobiol. Dis. 2000;7:623–643. doi: 10.1006/nbdi.2000.0299. [PubMed] [CrossRef] [Google Scholar]
68. Yang E.J., Jiang J.H., Lee S.M., Yang S.C., Hwang H.S., Lee M.S., Choi S.M. Bee venom attenuates neuroinflammatory events and extends survival in amyotrophic lateral sclerosis models. J. Neuroinflamm. 2010;7 doi: 10.1186/1742-2094-7-69. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
69. Cai M.D., Choi S.M., Yang E.J. The effects of bee venom acupuncture on the central nervous system and muscle in an animal hSOD1G93A mutant. Toxins. 2015;7:846–858. doi: 10.3390/toxins7030846. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
70. Jung G.B., Huh J.-E., Lee H.J., Kim D., Lee G.J., Park H.K., Lee J.D. Anti-cancer effect of bee venom on human MDA-MB-231 breast cancer cells using Raman spectroscopy. Biomed. Opt. Express. 2018;9:5703–5718. doi: 10.1364/BOE.9.005703. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
71. Lim H.N., Baek S.B., Jung H.J. Bee venom and its peptide component melittin suppress growth and migration of Melanoma Cells via inhibition of PI3K/AKT/mTOR and MAPK pathways. Molecules. 2019;24:929. doi: 10.3390/molecules24050929. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
72. Hong J., Lu X., Deng Z., Xiao S., Yuan B., Yang X. How Melittin inserts into cell membrane: Conformational changes, inter-peptide cooperation, and disturbance on the membrane. Molecules. 2019;24:1775. doi: 10.3390/molecules24091775. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
73. Liu S., Yu M., He Y., Xiao L., Wang F., Song C., Sun S., Ling C., Xu Z. Melittin prevents liver cancer cell metastasis through inhibition of the Rac1-dependent pathway. Hepatology. 2008;47:1964–1973. doi: 10.1002/hep.22240. [PubMed] [CrossRef] [Google Scholar]
74. Saidemberg D.M., Baptista-Saidemberg N.B., Palma M.S. Chemometric analysis of Hymenoptera toxins and defensins: A model for predicting the biological activity of novel peptides from venoms and hemolymph. Peptides. 2011;32:1924–1933. doi: 10.1016/j.peptides.2011.08.001. [PubMed] [CrossRef] [Google Scholar]
75. Liu X., Chen D., Xie L., Zhang R. Effect of honey bee venom on proliferation of K1735M2 mouse melanoma cells in-vitro and growth of murine B16 melanomas in-vivo. J. Pharm Pharmacol. 2002;54:1083–1089. doi: 10.1211/002235702320266235. [PubMed] [CrossRef] [Google Scholar]
76. Wang C., Chen T., Zhang N., Yang M., Li B., Lü X., Cao X., Ling C. Melittin, a Major Component of Bee Venom, Sensitizes Human Hepatocellular Carcinoma Cells to Tumor Necrosis Factor-related Apoptosis-inducing Ligand (TRAIL)-induced. Apoptosis by Activating CaMKII-TAK1-JNK/p38 and Inhibiting IkB alpha Kinase-NFkB. J. Biol Chem. 2009;284:3804–3813. doi: 10.1074/jbc.M807191200. [PubMed] [CrossRef] [Google Scholar]
77. Zheng J., Lee H.L., Ham Y.W., Song H.S., Song M.J., Hong J.T. Anti-cancer effect of bee venom on colon cancer cell growth by activation of death receptors and inhibition of nuclear factor kappa B. Oncotarget. 2015;6:44437–44451. doi: 10.18632/oncotarget.6295. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
78. Zuazo-Gaztelu I., Casanovas O. Unraveling the role of angiogenesis in cancer ecosystems. Front. Oncol. 2018;8 doi: 10.3389/fonc.2018.00248. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Rajabi M., Mousa S.A. The role of angiogenesis in cancer treatment. Biomedicines. 2017;5:34. doi: 10.3390/biomedicines5020034. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
80. Gülmez Y., Aydın A., Can I., Tekin S., Cacan E. Cellular toxicity and biological activities of honey bee (Apis mellifera L.) venom. Marmara Pharm. J. 2017;21:51–260. [Google Scholar]
81. Soman N.R., Baldwin S.L., Hu G., Marsh J.N., Lanza G.M., Heuser J.E., Arbeit J.M., Wickline S.A., Schlesinger P.H. Molecularly targeted nanocarriers deliver the cytolytic peptide melittin specifically to tumor cells in mice, reducing tumor growth. J. Clin. Investig. 2009;119:2830–2842. doi: 10.1172/JCI38842. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
82. Cheng B., Thapa B., Remant K.C., Xu P. Dual secured nano-melittin for the safe and effective eradication of cancer cells. J. Mater. Chemb. 2015;3:25–29. doi: 10.1039/C4TB01401D. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
83. Jallouk A.P., Moley K.H., Omurtag K., Hu G., Lanza G.M., Wickline S.A., Hood J.L. Nanoparticle Incorporation of Melittin Reduces Sperm and Vaginal Epithelium Cytotoxicity. PLoS ONE. 2014:9. doi: 10.1371/journal.pone.0095411. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
84. Hegazi A., Abdou A.M., EI-Moez S.I., Abd Allah F. Evaluation of the antibacterial activity of bee venom from different sources. World Appl. Sci. J. 2014;30:266–270. [Google Scholar]
85. Socarras K.M.T., Theophilus P.A.S., Torres J.P., Gupta K., Sapi E. Antimicrobial Activity of Bee Venom and Melittin against Borrelia burgdorferi. Antibiot. (Basel) 2017;6 doi: 10.3390/antibiotics6040031. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
86. Perumal Samy R., Gopalakrishnakone P., Thwin M.M., Chow T.K., Bow H., Yap E.H., Thong T.W. Antibacterial activity of snake, scorpion and bee venoms: A comparison with purified venom phospholipase A2 enzymes. J. Appl Microbiol. 2007;102:650–659. doi: 10.1111/j.1365-2672.2006.03161.x. [PubMed] [CrossRef] [Google Scholar]
87. Zolfagharian H., Babaie M. Antimicrobial activity of bee venom and Melittin against Borrelia burgdorferi. J. Pharmacopuncture. 2016;19:225–230. doi: 10.3831/KPI.2016.19.023. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
88. Uddin M.B., Lee B.H., Nikapitiya C., Kim J.H., Kim T.H., Lee H.C., Kim C.G., Lee J.S., Kim C.J. Inhibitory effects of bee venom and its components against viruses in vitro and in vivo. J. Microbiol. 2016;54:853–866. doi: 10.1007/s12275-016-6376-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
89. Bachis A., Cruz M.I., Mocchetti I. M-tropic HIV envelope protein gp120 exhibits a different neuropathological profile than T-tropic gp120 in rat striatum. Eur. J. Neurosci. 2010;32 doi: 10.1111/j.1460-9568.2010.07325.x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
90. Cecilio A.B., Caldas S., Oliveira R.A., Santos A.S., Richardson M., Naumann G.B., Schneider F.S., Alvarenga V.G., Estevão-Costa M.I., Fuly A.L., et al. Molecular characterization of Lys49 and Asp49 phospholipases A₂from snake venom and their antiviral activities against Dengue virus. Toxins. 2013;5:1780–1798. doi: 10.3390/toxins5101780. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
91. Fenard D., Lambeau G., Valentin E., Lefebvre J.C., Lazdunski M., Doglio A. Secreted phospholipases A2, a new class of HIV inhibitors that block virus entry into host cells. J. Clin. Investig. 1999;104:611–618. doi: 10.1172/JCI6915. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
92. Fenard D., Lambeau G., Maurin T., Lefebvre J.C., Doglio A. A peptide derived from bee venom-secreted phospholipase A2 inhibits replication of T-cell tropic HIV-1 strains via interaction with the CXCR4 chemokine receptor. Mol. Pharmacol. 2001;60:341–347. doi: 10.1124/mol.60.2.341. [PubMed] [CrossRef] [Google Scholar]
Zdroj: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6720840/