

1.Infection Medicine, Dekanát biomedicínskych vied, College of Medicine and Veterinary Medicine, The University of Edinburgh, Edinburgh, Spojené kráľovstvo
2.School of Medicine, Univerzita Kabale, Kabale, Uganda
3.Katedra biosystémového inžinierstva, Fakulta environmentálneho inžinierstva a strojného inžinierstva, Poznaňská univerzita, Poznaň, Poľsko
4.Katedra farmaceutickej chémie, College of Pharmacy, Univerzita Taif, Taif, Saudská Arábia
5.Fakulta poľnohospodárstva a živočíšnych vied, Busitema University Arapai Campus, Soroti, Uganda
6.Fakulta biomedicínskych vied, Kampala International University Western Campus, Bushenyi, Uganda
7.Katedra klinickej farmácie a farmaceutickej praxe, School of Pharmacy, Kampala International University Western Campus, Bushenyi, Uganda
8.Katedra lekárskej mikrobiológie a imunológie, Lekárska fakulta, Assiut University, Assiut, Egypt
9.Katedra základných lekárskych vied, Michael Chilufya Sata School of Medicine, Copperbelt University, Ndola, Zambia
10.Katedra farmakológie a terapie, Fakulta veterinárneho lekárstva, Univerzita Damanhour, Damanhour, Egypt
11.Národné výskumné centrum pre choroby prvokov, Univerzita poľnohospodárstva a veterinárnej medicíny Obihiro, Obihiro, Japonsko
12.Zhejiang University-University of Edinburgh Institute, Zhejiang University School of Medicine, Zhejiang University, Haining, Čína
Ťažký akútny respiračný syndróm koronavírus 2 (SARS-CoV-2) je charakterizovaný syndrómom závažnej cytokínovej búrky po zápale. SARS-CoV-2 priamo interaguje s receptormi angiotenzín-konvertujúceho enzýmu 2 (ACE-2) v ľudskom tele. Doplnkové terapie, ktoré ovplyvňujú expresiu IgE a IgG protilátok, vrátane podávania včelieho jedu (BV), sú účinné pri liečbe artritídy a Parkinsonovej choroby. Nedávna epidemiologická štúdia v Číne ukázala, že miestni včelári majú určitú úroveň imunity proti SARS-CoV-2 s predchádzajúcim vystavením vírusu aj bez neho. Protizápalové vlastnosti BV sú spojené s melitínom a fosfolipázou A2 (PLA2), pričom obe vykazujú aktivitu proti obaleným a neobaleným vírusom, vrátane H1N1 a HIV, s aktivitou sprostredkovanou antagonistickou aktivitou proti interleukínu-6 (IL-6), IL-8, interferón-y (IFN-y) a tumor nekrotizujúci faktor-a (TNF-a). Melitín je spojený s nedostatočnou expresiou prozápalových cytokínov, vrátane jadrového faktora-kappa B (NF-KB), kináz regulovaných extracelulárnym signálom (ERK1/2) a proteínkinázy Akt. BV terapia tiež zahŕňa sekrečnú fosfolipázu A2 skupiny III pri liečbe respiračných a neurologických ochorení. BV aktivácia bunkového a humorálneho imunitného systému by sa mala preskúmať na aplikáciu komplementárnej medicíny na zvládanie infekcií SARS-CoV-2. BV „vakcinácia“ sa používa na imunizáciu proti cytomegalovírusu a môže potlačiť metastázy prostredníctvom PLA2 a fosfatidylinozitol-(3,4)-bisfosfátových dráh. To, že BV vykazuje účinnosť na HIV a H1NI, ponúka príležitosť ako kandidáta na doplnkovú terapiu na ochranu pred SARS-CoV-2.
Úvod
Ťažký akútny respiračný syndróm koronavírus 2 (SARS-CoV-2) je pôvodcom koronavírusovej choroby 2019 (COVID-19), respiračnej infekcie, ktorá sa objavila v provincii Wuhan v Číne koncom roka 2019 (1, 2) a stala sa globálnou pandémiou v roku 2020. K 1. aprílu 2020 dosahovala celosvetová úmrtnosť 5 % (3). V priebehu niekoľkých týždňov sa celosvetová úmrtnosť zvýšila na 6,7 % (5 % v Afrike, 4,4 % v Amerike, 5 % v regióne východného Stredomoria, 4,4 % v juhovýchodnej Ázii, 8,9 % v európskom regióne, 4,4 % v západnej tichomorská oblasť) (4). Výzvy v oblasti verejného zdravia spôsobené ochorením COVID-19 sú obrovské, vrátane zvládania vysokého počtu asymptomatických prípadov (5). Choroba prehĺbila existujúce sociálno-ekonomické rozdiely, najmä v zraniteľných komunitách v rozvojových krajinách vrátane Afriky, ktoré boli neúmerne postihnuté dôsledkami extrémnych preventívnych opatrení (6).
Ťažké infekcie SARS-CoV-2 sú charakterizované syndrómom cytokínovej búrky, hyperzápalom a multiorgánovým zlyhaním (2, 7). Hostiteľské bunky sú infikované prostredníctvom receptora angiotenzín-konvertujúceho enzýmu 2 (ACE-2) (8, 9), ktorý je spojený s vrodenou aj získanou imunitou (10). Predpokladá sa, že ACE2 zvyšuje replikáciu vírusu a zosilňuje inváziu hostiteľských buniek (10) a je hlavnou zložkou systému renín-angiotenzín-aldosterón (RAAS), ktorý interaguje s enzýmami CVS a kaskáduje kardiovaskulárne ochorenia (11, 12). ACE2 môže byť dôvodom, prečo pacienti so SARS-CoV-2 vyžadujú farmakologickú profylaxiu trombózy (13, 14); patogenéza SARS-CoV-2 zahŕňa väzbu vírusu na epitelové bunky a lokálnu propagáciu s minimálnou vrodenou imunitnou odpoveďou (15). Druhé štádium infekcie vykazuje zvýšené šírenie vírusu, aktívnu imunitnú odpoveď, šírenie vírusu do dolného dýchacieho systému a môže zahŕňať kardiovaskulárny a tráviaci systém (16). Tretie štádium zahŕňa hypoxiu, infiltráciu celého dýchacieho systému a nakoniec syndróm akútnej respiračnej tiesne (ARDS), ktorý je potenciálne smrteľný (15). SARS-CoV-2 sa spája s koagulopatiami, trombotickými príhodami (17) a vyčerpaním lymfocytov (18).
V súčasnosti neexistuje žiadny celosvetovo uznávaný protokol alternatívnej liečby SARS-CoV-2, hoci podávanie polyklonálnych protilátok je sľubné (19). Účinnosť chlorochínu a jeho derivátov sa naďalej skúma pri prevencii COVID-19 (20, 21), ako aj famotidínu, protivredového lieku, podávaného vo vysokých dávkach (10× normálne) počas 14 dní na kontrolu SARS-CoV- 2 infekcia (7). Remdesivir, ktorý sa predtým používal na zvládnutie blízkovýchodného respiračného syndrómu-kronavírusu (MERS-CoV), bol skúmaný ako kandidátsky liek proti SARS-CoV-2 (22–24). Kombinácie Lopinavir/ritonavir, bežne používané na prevenciu HIV/AIDS, sa tiež skúmajú z hľadiska účinnosti proti SARS-CoV-2 (25, 26). Neutrofilné extracelulárne pasce (NET), bežné u hadov, hmyzu, pavúkovcov a myriapodov, sa tiež zvažovali pre SARS-CoV-2 (27, 28). Včelí jed (BV) môže pôsobiť ako inhibítory ACE2 alebo blokátory angiotenzínových receptorov (ARB), hoci štúdie o BV a SARS-CoV-2 sú zriedkavé. Je známe, že hadí jed pôsobí prostredníctvom fosfolipázy A2 (PLA2) na produkciu kyseliny arachidónovej, ktorá vyvoláva hypotenziu (29). U ľudí jed blanokrídlovcov znížil kľúčové parametre v RAAS (30). Kombinácia BV a propolisu bola spojená s hypotenziou u laboratórnych zvierat prostredníctvom zníženia hladín angiotenzínu v sére (31), čo dokazuje úzky vzťah medzi BV a dráhou ACE2.
Terapia včelím jedom
Terapia včelím jedom (BV) sa datuje do éry Hippokrata, kde bola nasadená na zmiernenie bolesti kĺbov a artritídy (32). V súčasnej medicíne sa BV používa na liečbu roztrúsenej sklerózy (33), artritídy a Parkinsonovej choroby (34). Aktivita je založená na výhodách anafylaktickej reakcie na metabolizmus a na organely, najmä v dýchacom systéme (35). Alergény môžu ponúkať výhody proti COVID-19 (36, 37); BV môže vyvolať zvýšenie špecifických IgE a IgG protilátok (38) a vedie k produkcii IgE protilátok, ktoré môžu reagovať na rôzne antigény (39) (tabuľka 1). Hoci IgE sú zodpovedné za alergické vzplanutia, ponúkajú aj ochrannú úlohu hostiteľa v rámci širokého spektra alergénov (39). BV môže pôsobiť ako adjuvans, keď sa kombinuje s Toll-like receptormi (TLR) ligandami (40) a moduluje imunitný systém, zvyšuje diferenciáciu buniek exprimujúcich foxP3 a zvyšuje cirkulujúce regulačné T bunky (41, 42). BV spúšťa zvýšenie CD25, CD4+ T buniek a foxP3 mRNA, čo vedie k posunu v BV-špecifickom pomere IgG4/IgE (43). BV reguluje imunitnú odpoveď a fyziopatologické zmeny (44) a podporuje klinické pozorovania pri apiterapii, kde sa včelárom ukázalo, že zvyšujú imunitu proti COVID-19 v provincii Wuhan, PR Čína (45).
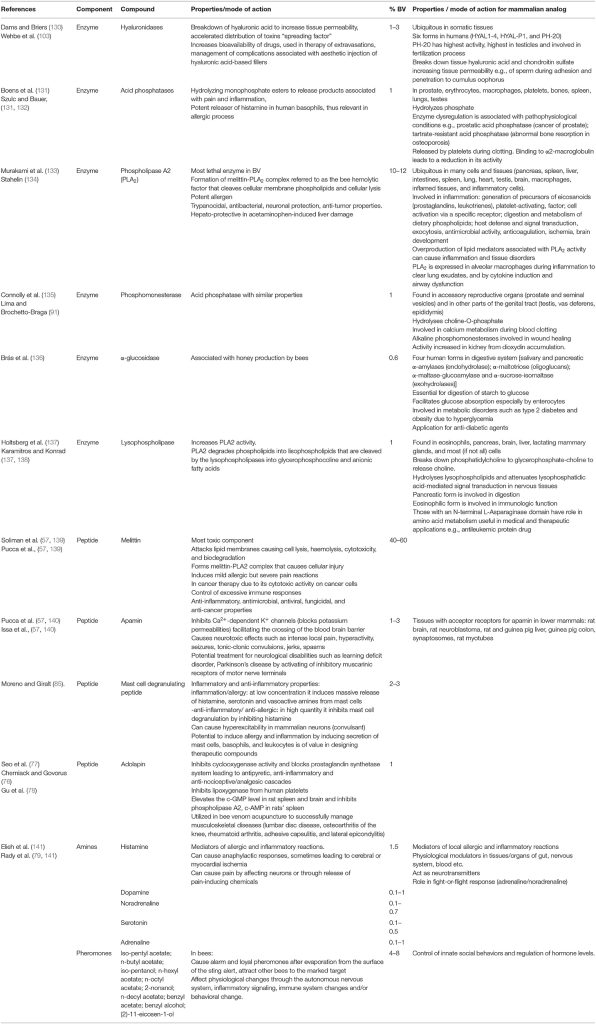
bvPLA2 môže spustiť dozrievanie žírnych buniek (46), je dôležitý pri bunkovej signalizácii a pri produkcii kľúčových lipidov a môže pôsobiť ako receptorový ligand (47). PLA2 môže inhibovať tok zápalových buniek k cieľom (48). BV môžu viesť k trvalej indukovanej tolerancii na príbuzné alergény (49), ako funkcia zníženia IgG4 a aktivácie IL-10, modulácie imunitného systému a indukcie odchýlky od TH2 k TH1 (50–52). Melittín (APi M 1) sa môže použiť na vývoj mimotopov (49). APi M 10 (icarapin), zložka BV, aktivuje efektorové bunky pacientov alergických na včelí jed (53). Keďže IgE má epitop pre APi M 10, môže to ponúknuť príležitosť na vývoj adjuvans. Antigény BV možno použiť ako adjuvans pri liečbe bolesti (54) a pôsobenie melitínu na tvorbu pórov bunkovej membrány (54, 55), čo vedie k apoptóze, slúži na posilnenie adjuvantných vlastností. BV má tiež antivírusové vlastnosti (56). BV môže znecitlivieť žírne bunky a bazofily (57) a potlačiť vrodené lymfoidné bunky. Materiály BV môžu inhibovať syntézu proteínov, indukovať angiogenézu (58) a aktivovať kaspázu-3-8-9 (59) (tabuľka 1).
Podmienky, ktoré umožňujú použitie včelieho jedu napriek jeho toxicite
Včelí jed je vo vysokých dávkach cytotoxický, avšak necytotoxické koncentrácie BV sa pohybujú od 1 do 3 μg/ml a vykazujú významný terapeutický potenciál (60). Nízke dávky, kontrolované koncentrácie a zriedené BV spúšťajú celý rad protizápalových reakcií (61, 62) a boli nasadené na liečbu cukrovky, reumatoidnej artritídy (RA), srdcových chorôb, obezity, astmy, kožných chorôb a centrálnej choroby spojené s nervovým systémom, ako je Alzheimerova choroba, Parkinsonova choroba a skleróza (61–64). V nízkych dávkach môže BV potlačiť zápalové cytokíny, ako je interleukín-6 (IL-6), IL-8, interferón-y (IFN-y) a tumor nekrotizujúci faktor-a (TNF-a). Zníženie signálnych dráh zodpovedných za aktiváciu zápalových cytokínov, ako je nukleárny faktor-kappa B (NF-κB), kinázy regulované extracelulárnym signálom (ERK1/2) a proteínkináza Akt a lipopolysacharid porfyromonas gingivalis (PgLPS)- liečených ľudských keratinocytov bola spojená s liečbou zahŕňajúcou BV (65).
BV sa používa ako protizápalové činidlo kombináciou zlúčenín v BV, t.j. sekrečnej fosfolipázy A2, s fosfatidylinozitol-(3,4)-bisfosfátom alebo bunkami, najmä dendritickými bunkami (DC), alebo kombináciou BV s DC (66) . Konjugácia hormonálnych receptorov a transportérov génovej terapie k BV peptidom ako užitočná nová cielená terapia na pozitívnu moduláciu imunitných odpovedí bola aplikovaná v protirakovinovej a protizápalovej terapii (67).
Imunitné reakcie BV sú pri vysokých dávkach toxické, ale keď sú kontrolované alebo zriedené (kontrolované koncentrácie), tieto imunitné reakcie môžu slúžiť ako imunitné modulátory. Kontrolovaná alergická imunita môže byť výhodná pre obranu hostiteľa proti antigénom a patogénom vrátane RNA vírusov. BV môže stimulovať imunitné reakcie typu 2, imunita typu 2 je iniciovaná protilátkami T-buniek (T-pomocník typu 2) a imunoglobulínových (Ig) protilátok (IgE a IgG1) a pôsobením vrodeného imunitného systému, ako je epitel a biela krviniek a slúži ako bariérová obrana na elimináciu antigénov (68). BV skupina III sPLA2 vykazuje in vitro a in vivo účinky na imunitný systém. Modulované imunitné reakcie z BV môžu zmierniť imunologické ochorenia, ako je reumatoidná artritída, zápalové ochorenia, astma a Parkinsonova choroba (69). Vrodený imunitný systém indukuje obrannú imunitnú odpoveď proti antigénom BV prostredníctvom receptorov rozpoznávania vzorov (PRR), vrátane receptorov podobných Toll, ktoré sa nachádzajú na molekulárnych vzoroch spojených s patogénmi (PAMP) (70). BV pri terapeutickom ochorení je protizápalový (44), znižuje počet infiltrovaných zápalových buniek a expresiu tumor nekrotizujúceho faktora (TNF)-a, interleukínu (IL)-1p, inhibíciu Toll-like receptora (TLR)2 a CD14 . BV tiež potláčajú väzbový potenciál jadrového faktora-κB (NF-κB) a aktivačného proteínu (AP)-1 (71). Ľudský receptor IL-1 (anakinra) tiež vykazuje protizápalovú aktivitu (72), avšak informácie spájajúce tento receptor a včelí jed sú riedke.
Fosfolipáza 2 z včelieho jedu (bvPLA2) je hlavným alergénom v BV a stimuluje vrodený imunitný systém väzbou na receptory rozpoznávajúce vzory (PRR), napr. 2 imunitná odpoveď. bvPLA2 indukuje reakcie typu T-helper buniek a vrodené lymfoidné bunky skupiny 2 (ILC2) uľahčené enzýmom podporovaným štiepením membránových fosfolipidov a sekréciou IL-33. bvPLA2 tiež indukuje produkciu IgE, o ktorej sa ukázalo, že má u myší ochranu pred budúcimi alergickými/imunologickými reakciami [v prípade letálnej dávky BV (70)]. PLA2 hrá dôležitú úlohu v obrane hostiteľa pri diferenciácii Th2, aktivácii ILC2, produkcii imunoglobulínov, prestavbe membrány a protizápalových reakciách (44, 70).
BV vykazuje pozitívne imunomodulačné úlohy; zníženie progresie nádorov a aktivácia imunitného systému kombináciou bvPLA2 s fosfatidylinozitol-(3,4)-bisfosfátom alebo bunkami, najmä dendritickými bunkami (DC) (66). DC pripravené s BV in vivo majú protirakovinové aj antivírusové vlastnosti. DC kombinované s antigénmi z nádoru alebo vírusu produkujú peptidové epitopy hlavného histokompatibilného komplexu (MHC) triedy I a II pre CD8 a CD4 T lymfocyty (obrázok 1).

PLA2 (bvPLA2-H34Q) sa viaže na membránu a in vivo kombinuje antigény s membránou ľudských DC buniek, čo spôsobuje stimuláciu CD8 T buniek a antivírusové a protinádorové vakcíny (DC vakcína) možno získať od BV pomocou DC. Tieto antivírusové/protinádorové vakcíny na báze buniek sa používajú počas imunizácie proti vírusom vrátane cytomegalovírusu a na potlačenie nádorov (73, 74). BV je známa antimikrobiálna a protinádorová vakcína potencovaná adjuvans. Melittín, bvPLA2 a fosfatidylinozitol-(3,4)-bisfosfát sú účinnými adjuvans pre vakcíny proti leishmánii, proti nádorom a proti cytomegalovírusom (73–75). Konjugácia BV peptidov s hormonálnymi receptormi a génová terapia ponúkajú pozitívnu moduláciu imunitných reakcií aplikované ponúkajú cielené protirakovinové a protizápalové terapie (67).
BV sa môže použiť ako analgetikum v kontrolovaných koncentráciách dávky; inhibícia aktivity cyklooxygenázy a blokovanie systému prostaglandínsyntetázy, čo vedie k antipyretickým, protizápalovým a antinociceptívnym/analgetickým kaskádam (76–78). V zriedenej forme môže BV indukovať antinociceptívne účinky prostredníctvom α-adrenergného receptora (aktivácia miechového α-adrenergného receptora) (61, 62). Konjugácia BV peptidov s proteínovými receptormi, ako sú hormóny a gény transportujúce peptidy, poskytuje inovatívnu BV kontrolovanú protizápalovú, antinociceptívnu a imunomodulačnú terapiu (67).
Farmakodynamika zložiek včelieho jedu
Včelí jed (BV) obsahuje enzýmy [fosfolipáza A2 (PLA2), fosfolipáza B, hyaluronidázy, kyslé fosfatázy, kyslé fosfomonesterázy, α-D-glukozidázy a lyzofosfolipázy]; peptidy lytický peptid melitín, apamín, mastocyty (žírne bunky) degranulujúci peptid, sekapín, pamín, minimín, procamín A, B, inhibítor proteázy, tertiapín, kardiopep a adolapín; a aminokyseliny zahŕňajú kyselinu g-aminomaslovú a a-aminokyseliny. Nepeptidové zložky zahŕňajú amíny (dopamín, histamín, norepinefrín, neurotransmitery), sacharidy (glukóza, fruktóza), feromóny (izo-pentylacetát; n-butylacetát; izo-pentanol; n-hexylacetát; n-oktylacetát; 2 -nonanol, n-decylacetát, benzylacetát, benzylalkohol a (2)-11-eikosen-1-ol) (79, 80) (tabuľka 1).
Ukázalo sa, že BV má protizápalové, antinociceptívne, antioxidačné a antiapoptotické vlastnosti a ukázalo sa, že mení génovú expresiu a fibrózu (81–84). Vedľajšie účinky zahŕňajú prozápaly [vyššie dávky PLA2, peptidy degranulujúce žírne bunky, hemolytické zlúčeniny (melittín)], alergické reakcie na inhibítory proteázy a peptidy, anafylaktické reakcie a smrť (76).
Za alergickú reakciu sú zodpovedné viaceré proteínové alergény vo včelom jede (85). Alergické reakcie sa môžu vyskytnúť v dýchacom systéme, gastrointestinálnom systéme, kardiovaskulárnom systéme, koži a bodnutí a môžu vyústiť do ťažkého anafylaktického šoku, niekedy vedúceho k cerebrálnej alebo myokardiálnej ischémii (86, 87). Imunitne nesprostredkovaný mechanizmus alergie na BV zahŕňa produkciu mediátorov bradykinínu (BK), čo vedie k anafylaxii (88) z melitínovej aktivácie PLA2 (napodobňovanie BK).
Biologická variabilita zloženia včelieho jedu medzi variantmi včiel na podávanie biotoxínov v komplementárnej medicíne
Včely a osy patria do radu hmyzu Hymenoptera (89, 90). U včiel je produkcia jedu najvyššia u včelích kráľovien pri vzchádzaní. Jed blanokrídlovcov spôsobuje toxické alebo alergické reakcie väčšinou spôsobené biochemickými zlúčeninami spojenými s lokálnym zápalovým pôsobením (91, 92). Žihadlá bránia kolóniu všetkých druhov hmyzu radu Hymenoptera (93, 94). Melittín je najvýznamnejšou zlúčeninou zodpovednou za tieto alergické reakcie (95, 96); hoci kombinácia mastocytov s IgE vyvoláva aktivitu leukotriénov, histamínov a faktorov aktivujúcich krvné doštičky počas alergických reakcií (93, 94, 97).
Jedy blanokrídlovcov obsahujú dopamín, adrenalín, hyaluronidázu, noradrenalín, serotonín, histamín, fosfolipázy A a B (85), ale iba BV obsahuje peptid degranulujúci žírne bunky, melitín a apamín (57). Rôzne druhy včiel; Apis mellifera mellifera a Apis mellifera ligustica (v Európe) a Apis mellifera scutellate (v Afrike) sú zodpovedné za ľudské otravy (57). Stredná letálna dávka BV sa pohybuje od 2,8 do 3,5 mg/kg telesnej hmotnosti a pri epizóde bodnutia sa v priemere podá 140–150 μg BV (57). Pravdepodobnosť úmrtia len na niekoľko bodnutí včelou je u nealergických osôb minimálna (98) a závažnosť otravy je náležite ovplyvnená telesnou hmotnosťou, vekom a imunitným stavom obete (99, 100). Číslo bodnutia a akákoľvek predchádzajúca senzibilizácia na BV ovplyvňuje závažnosť otravy (99, 100).
BV je číra, bezfarebná vodnatá kvapalina bez zápachu s pH 4,5–5,5 s horkastou chuťou a v niektorých prípadoch s okrasným štipľavým zápachom (101, 102). Zloženie BV je ovplyvnené extrakčnými metódami kvôli jeho prchavosti (101). Jed Apis mellifera je pravdepodobne najlepšie charakterizovaným jedom blanokrídlovcov (103). Jed zo všetkých druhov Apis je podobný zložením a kvalitou. A. florea, včela medonosná, je najmenšia vo svojej rodine, zatiaľ čo A. dorsata je najväčšia (101). Apis cerana jed je dvakrát tokický ako jed Apis mellifera (104). V jedových žľazách A. dorsata > A. cerana > A. mellifera > A. florea boli pozorované rozdiely v zložení sekrécie jedovej žľazy a jedového vaku a koncentrácii lipidov, proteínov, aktivite kyslej fosfatázy a hexokinázy. Zistilo sa, že kompozície lipidov, proteínov, sacharidov a alkalickej fosfatázy sú v poradí A. cerana > A. mellifera > A. florea. Glykogén chýbal v jedovej žľaze aj v jedovom vaku druhu Apis (101).
Variabilita v zložení včelieho jedu súvisí s druhom, vekom, geografickou lokalizáciou a sociálnym stavom (96). Mladé včely robotnice majú nižšie koncentrácie melitínu a histamínu a vyššie koncentrácie apamínu ako staršie včely robotnice (57). Včelie kráľovné majú nízke koncentrácie melitínu a apamínu a vysoké koncentrácie histamínu (57). APi M dosahuje svoj vrchol, keď má včela ~28 dní a s vekom klesá (105). Hladiny PLA2 dosahujú maximum okolo 10. dňa vyliahnutia (101). Africké včely uvoľňujú malé množstvá jedu pri bodnutí, s vysokými koncentráciami PLA2 a zníženými koncentráciami melitínu a hyaluronidázy (57). Boli hlásené sezónne odchýlky v zložení BV (106); napríklad počas zimy sa produkcia APi M zvyšuje, ale počas leta klesá (107, 108).
Súčasné terapeutické pokroky včelieho jedu
Antivírusové a antibakteriálne vlastnosti
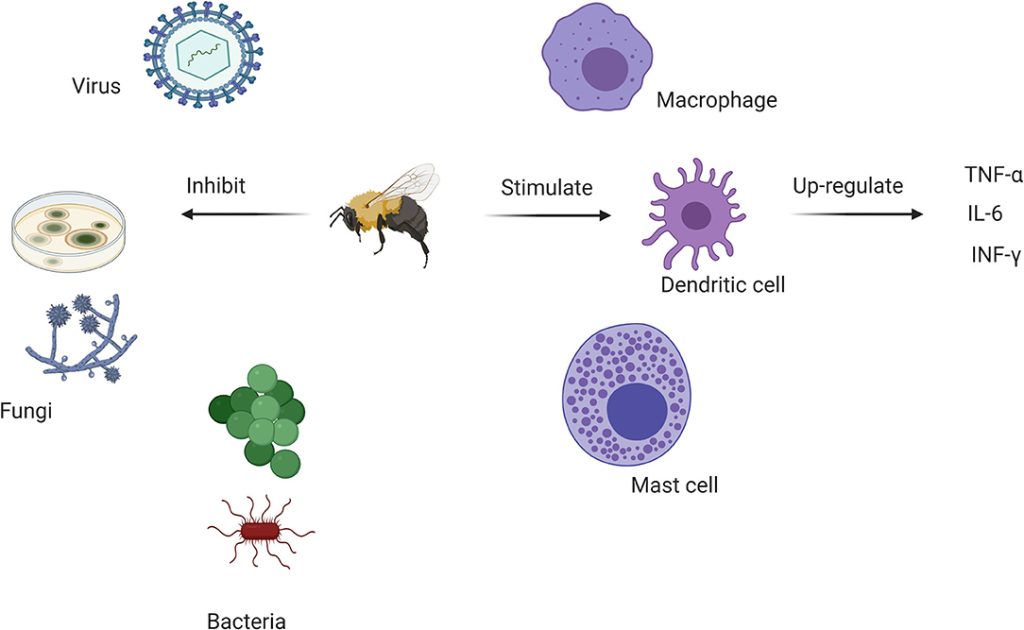
Melittín a PLA2 vykazujú antimikrobiálne aktivity a boli použité ako doplnkové antibakteriálne činidlá (103); vyvolanie tvorby pórov a deštrukcie baktérií (109). APi M vykazuje antivírusové vlastnosti proti niektorým obaleným vírusom a neobaleným vírusom in vitro (110). Ochrana bola pozorovaná u myší po expozícii vírusu chrípky A H1N1, ale BV môže tiež interagovať priamo s vírusovým povrchom (110) (obrázok 2).
Manažment rakoviny
BV bol skúmaný pri rakovine (111, 112); melitín sa považuje za cytolytický, ale nešpecifický. Melittín môže rozložiť membránovú lipidovú dvojvrstvu a pri intravenóznej injekcii vykazuje toxicitu (113). APi M má schopnosť potlačiť rast nádorov v bunkách rakoviny prsníka, pečene, prostaty a pľúc (111, 112). Štúdie in vitro a in vivo ukazujú, že melitín môže potlačiť rast rakovinových buniek inhibíciou signalizácie NF-KB a aktiváciou dráh kaspázy 3 a 9. Inhibícia motility buniek hepatocelulárneho karcinómu sa pozorovala in vitro a in vivo potlačením dráh závislých od Rac1 (114).
Protizápalový potenciál
Nízke dávky BV spúšťajú celý rad protizápalových reakcií, ktoré sa skúmali pri cukrovke, reumatoidnej artritíde (RA), srdcových ochoreniach, obezite, astme, kožných ochoreniach a ochoreniach spojených s centrálnym nervovým systémom (Alzheimerova choroba, Parkinsonova choroba a amyotrofická laterálna skleróza) (63, 64). BV potláča zápalové cytokíny, vrátane interleukínu-6 (IL-6), IL-8, interferónu-y (IFN-y) a tumor nekrotizujúceho faktora-a (TNF-a). Zníženie signálnych dráh zodpovedných za aktiváciu zápalových cytokínov, nukleárneho faktora-kappa B (NF-KB), kináz regulovaných extracelulárnym signálom (ERK1/2) a proteínkinázy Akt a lipopolysacharidu Porphyromonas gingivalis (PgLPS) liečených ľudí keratinocyty sú spojené s liečbou melitínom(65) (obrázok 2).
Odpovede hostiteľa na včelí jed /BV/
BV terapia môže zmierniť ochorenia súvisiace s imunitou. Sekrečná fosfolipáza A2 skupiny III z BV (BV skupina III sPLA2) vykazuje in vitro a in vivo aktivitu na imunitný systém a používa sa na zvládnutie astmy, Parkinsonovej choroby a zápalu orgánov vyvolaného liekmi (69). BV imunitné reakcie môžu byť nebezpečné, keď sú vysoko zvýšené, ale keď je kontrolovaná, alergická imunita môže byť výhodná pri obrane hostiteľa na stimuláciu imunitných reakcií 2. typu. Imunita 2. typu je založená najmä na bariérovej obrane a tieto reakcie sú iniciované pomocnými T bunkami typu 2 (TH2), protilátkami imunoglobulínov E a G1 (IgE a IgG1) a ďalšími zložkami vrodeného imunitného systému (epiteliálne bariéry, vrodené lymfoidné bunky-ILC, eozinofily, žírne bunky, bazofily a aktivované makrofágy) (68). Vrodený imunitný systém vníma zložky jedu a vyvoláva obrannú imunitnú odpoveď proti antigénom prostredníctvom receptorov rozpoznávania vzorov (PRR), napr. Toll-like receptorov, ktoré sa nachádzajú na molekulových vzorcoch spojených s patogénmi (PAMP) (70). Protizápalové vlastnosti BV (44) môžu inhibovať aktivitu zápalových antigénov, znížiť počet infiltrovaných zápalových buniek a inhibovať expresiu (TNF)-α, IL-1β, Toll-like receptor (TLR)2 a CD14 potláčajúc väzbovú aktivitu jadrového faktora-KB (NF-KB) a aktivačného proteínu (AP)-1 (71). Hlavný alergén Bet V 1, PLA2, stimuluje vrodený imunitný systém, viaže sa na PRR, napr. Toll-like receptory, ktoré rozpoznávajú PAMP, čím spúšťa imunitnú odpoveď typu 2 u myší. PLA2 v BV indukuje reakcie typu T pomocných buniek typu 2 (Th2) a aktiváciu vrodených lymfoidných buniek skupiny 2 (ILC2) prostredníctvom enzymatického štiepenia membránových fosfolipidov a sekrécie IL-33. PLA2 indukuje produkciu IgE, čím chráni myši pred budúcimi alergickými/imunologickými reakciami v prípade letálnej dávky BV (70); PLA2 hrá rozhodujúcu úlohu v obrane hostiteľa zlepšením diferenciácie Th2, aktivácie ILC2, produkcie imunoglobulínov, prestavby membrány a protizápalových reakcií (44, 70).
Vakcíny
BV môže potlačiť progresiu nádorov a aktivovať imunitný systém kombináciou sekrečnej fosfolipázy A2 v BV so zlúčeninami vrátane fosfatidylinozitol-(3,4)-bisfosfátu alebo dendritických buniek (DC) (66). DC ošetrené BV in vivo vykazujú protirakovinové a antivírusové vlastnosti. DC kombinované s antigénmi z nádoru alebo vírusu môžu produkovať peptidové epitopy hlavného histokompatibilného komplexu (MHC) triedy I a triedy II pre CD8 a CD4 T lymfocyty, čo vedie k sérii imunitných reakcií v reakcii na antigény. BV fosfolipáza A2 (bvPLA2-H34Q) sa viaže na membránu a spája antigény v bunkovej membráne ľudských DC in vivo. To indukuje rozpoznávanie a aktiváciu CD8 T buniek s tým, že antivírusové a protinádorové vakcíny môžu byť odvodené z BV (DC vakcína). Vakcíny od BV a DC (bunkové antivírusové/protinádorové vakcíny) sa používajú na imunizáciu proti vírusom, ako je cytomegalovírus, a na potlačenie nádorov (73, 74). BV možno použiť ako silnú antimikrobiálnu a protinádorovú vakcínu zosilnenú adjuvans a vykazuje potenciál vo vakcínach obsahujúcich melitín. sPLA2 a fosfatidylinozitol-(3,4)-bisfosfát sú účinnými adjuvans (proti leishmánii, protinádorovým a anticytomegalovírusovým vakcínam) (73–75).
Vedúcim adjuvantom terapií SARS-CoV-2, ktoré sa v súčasnosti podporuje, je hydroxid hlinitý kvôli jeho pomalému uvoľňovaniu a zvýšenej interakcii s bunkami prezentujúcimi antigén (115). Včelí jed ponúka kandidáta na kontrolu infekcií SARS-CoV-2 a mohol by ponúknuť výhody proti COVID-19. PLA2 bola spojená s úrovňou úspechu proti infekciám SARS-CoV-2 (116, 117). Konjugácia BV peptidov by mohla ponúknuť nový prístup vo vývoji BV vakcíny.
Potenciálny vzťah medzi proteínmi včelieho jedu a proteínmi COVID-19
SARS-CoV-2 patrí do rodu β-koronavírusov. SARS-CoV-2 má štyri zrejmé štrukturálne proteíny: membránový, hrotový, nukleokapsidový proteín a obal. Štrukturálna integrita vírusu SARS-CoV-2 je udržiavaná štrukturálnymi proteínmi a vytvára ochranný obal okolo jeho RNA. Membrána koronavírusu obsahuje 3 alebo 4 vírusové proteíny (118, 119), membránový glykoproteín je najhojnejším štrukturálnym proteínom a trikrát pokrýva membránovú dvojvrstvu, s dlhým COOH koncom vo virióne a krátkou NH2-koncovou doménou mimo vírusu (120). Genóm SARS-CoV-2 kóduje niekoľko čítacích rámcov (ORF); ORF1a/b kóduje 16 neštrukturálnych proteínov a prekladá dva polyproteíny (pp1a a pp1ab), ktoré tvoria až 2/3 vírusovej RNA. Zvyšné ORF kódujú štrukturálne proteíny (špičkový glykoproteín, matricový proteín, nukleokapsidový proteín a malý obalový proteín) (118, 119). SARS-CoV-2 má doplnkové proteíny, ktoré interferujú s vrodenou imunitnou odpoveďou hostiteľa (118).
Špičkový proteín je zvyčajne membránový glykoproteín typu I a tvorí peploméry, o ktorých je známe, že sa podieľajú na interakcii protilátky. Membrána hrá významnú úlohu pri intracelulárnej tvorbe vírusových častíc nezávisle od vírusovej špičky. Koronavírusy rastú a produkujú formy bez hrotov v prítomnosti tunicamycínu, čo vedie k produkcii neinfekčných viriónov, ktoré obsahujú membrány, ale bez hrotov (118).
Melittín môže prepichnúť ochranné membránové obaly obklopujúce vírusy vrátane vírusu ľudskej imunodeficiencie (HIV) (119). Mnohé vírusy, vrátane SARS-CoV-2, sa spoliehajú na svoj ochranný obal a môžu byť zraniteľné pri terapii včelím jedom vedeným melitínom (tabuľka 1).
Zložky fosfolipázy A2 včelieho jedu majú potenciál pre antivírusové aktivity (121). Nanočastice naplnené melittínom dodávali značné množstvo melitínu intravenózne, pričom sa zameriavali a zabíjali prekancerózne lézie u myší K14-HPV16 so skvamóznou dyspláziou a karcinómom obsahujúcim transgénne prvky ľudského papilomavírusu (HPV) (onkogény E6 a E7) (122, 123).
V provincii Chu-pej, epicentre prepuknutia SARS-CoV-2 v Číne, vykonalo miestne združenie včelárov medzi 23. februárom a 8. marcom prieskum medzi 5 115 včelármi (vrátane 723 vo Wu-chane) a ukázalo sa, že u žiadneho sa u pacientov s COVID-19 nerozvinuli žiadne príznaky ( 124). Opýtaných bolo päť apiterapeutov vo Wu-chane a 121 ich pacientov, ktorí absolvovali apiterapiu v období od októbra do decembra 2019; dvaja apiterapeuti boli vystavení podozrivým a/alebo potvrdeným obetiam COVID-19 bez ochrany. U žiadneho z apiterapeutov sa nerozvinuli symptómy súvisiace so SARS-CoV-2 a žiadny z ich 121 pacientov sa nenakazil COVID 19, napriek tomu, že 3 boli vystavení príbuzným infikovaným SARS-CoV-2 (124).
Apiterapia využíva včely medonosné a ich produkty (BV, med, materská kašička, peľ, propolis, včelí vosk). BV terapia využíva jed na moduláciu imunitného systému tela a zlepšenie/uľahčenie hojenia a zahŕňa buď použitie živých včelích bodnutí alebo injekčného jedu na liečbu artritídy, reumatoidnej artritídy, roztrúsenej sklerózy (MS), lupusu, ischias, bolesti krížov, a tenisový lakeť (125, 126). Produkty blanokrídlovcov sú silnými urýchľovačmi hojenia rán (127). Hmyzí jed je v porovnaní s hadím jedom menej zložitý a menej variabilný v zložení a fyziologickej aktivite (125, 126). BV sa môže podávať na vyvolanie alergických imunitných reakcií stimulujúcich vrodený imunitný systém hostiteľa (68) v dôsledku prítomnosti alergénov, ktoré podporujú imunitné reakcie typu 2 (44, 68–71). Protivírusový a protinádorový účinok BV, keď sa BV sekrečná fosfolipáza A2 zmieša s inými zlúčeninami, ako je fosfatidylinozitol-(3,4)-bisfosfát alebo dendritické bunky a/alebo včelie proteíny, ako je melitín, je výhodný (66) a používa sa v produkcia bunkových antivírusových/protinádorových vakcín (73–75). Imunologické vlastnosti BV sa nachádzajú aj v prírodných produktoch, ktoré napodobňujú včelí jed (127, 128), a ďalších štúdiách týkajúcich sa úlohy včelieho jedu ako potenciálneho kandidáta na použitie v komplementárnej medicíne pri liečbe vírusov, ako je SARS-CoV- 2 mohli zvážiť iné prírodné produkty, ktoré napodobňujú aktivitu BV.
Budúci výskum včelieho jedu
Vývoj adjuvantných terapií (pomocou APi M a PLA2) na použitie proti infekciám SARS-CoV-2 ponúka jedinečný prístup k vírusovej terapii. Vývoj vakcíny proti včelímu jedu pomocou DC s použitím APi M a bvPLA2 ponúka novú príležitosť na doplnkové medicínske zásahy proti infekciám SARS-CoV-2. Štúdie na skúmanie bunkovej signalizácie medzi proteínmi BV, Janus kinázou (JAK) a aktivátorom transkripcie (JAK-STAT) by pomohli posilniť jej prijatie v komplementárnej medicíne proti SARS-CoV-2. Inhibítory JAK sú spojené so zlepšenou prognózou u pacientov s COVID-19 (72, 129), ale na objasnenie bunkových mechanizmov sú potrebné štúdie. Synergická aktivita prostredníctvom kombinácií v alternatívnej a komplementárnej medicíne by pomohla bojovať proti vedľajším účinkom spojeným so súčasnými monoterapiami na zvládanie infekcií SARS-CoV-2. SARS-CoV-2 je nový vírus a môžu byť potrebné nové terapie na podporu manažmentu v priebehu času a môžu byť cenné pri podpore imunitnej odpovede u pacientov trpiacich takzvaným dlhotrvajúcim COVID.
Záver
Účinky SARS-CoV-2 na receptory ACE2 sú spojené so závažnou zápalovou aktivitou a zlou prognózou v závislosti od sprievodných ochorení pacienta a iných súvisiacich rizikových faktorov. Aj keď sa pacient zotaví z počiatočnej infekcie, môže byť konfrontovaný s dlhou a komplikovanou rekonvalescenciou a/alebo takzvaným dlhodobým ochorením COVID. Je nepravdepodobné, že čoskoro bude k dispozícii magická terapia COVID-19 a mali by sa preskúmať doplnkové terapie, ktoré dopĺňajú konvenčnú terapiu a podporujú zdravé zotavenie. BV melitín a aktivita fosfolipázy A2 majú silný protizápalový účinok a mohli by sa použiť na podporu zotavenia. To, že sa BV úspešne používa na zvládanie neurologických a imunologických ochorení, posilňuje dôvod na skúmanie jeho použitia v doplnkovej medicíne pri infekciách SARS-CoV-2. BV je potenciálnym adjuvans proti COVID-19, ktorý by sa mal pridať do zoznamu hlavných terapií.
Autorské príspevky
Všetci uvedení autori významne, priamo a intelektuálne prispeli k dielu a schválili ho na zverejnenie.
Financovanie
Túto prácu podporil Fond pre núdzový výskum nadácie Zhejiang University Education Foundation (SCW); Global Challenges Research Fund a University of Edinburgh.
Konflikt záujmov
Autori vyhlasujú, že výskum bol vykonaný bez akýchkoľvek obchodných alebo finančných vzťahov, ktoré by mohli byť chápané ako potenciálny konflikt záujmov.
Referencie
1. Ji Y, Ma Z, Peppelenbosch MP, Pan Q. Potential association between COVID-19 mortality and health-care resource availability. Lancet GlobHealth. (2020) 8:e480. doi: 10.1016/S2214-109X(20)30068-1
CrossRef Full Text | Google Scholar
2. Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ. COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet. (2020) 395:1033–4. doi: 10.1016/S0140-6736(20)30628-0
PubMed Abstract | CrossRef Full Text | Google Scholar
3. WHO. Coronavirus disease 2019. (COVID-19) Situation Report-72 HIGHLIGHTS. (2020). Available online at: https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200401-sitrep-72-covid-19.pdf?sfvrsn=3dd8971b_2 (accessed September 15, 2020).
4. WHO. Coronavirus disease 2019. (COVID-19) Situation Report – 88. (2020). Available online at: https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200417-sitrep-88-covid-191b6cccd94f8b4f219377bff55719a6ed.pdf?sfvrsn=ebe78315_6 (accessed September 15, 2020).
5. Mizumoto K, Kagaya K, Zarebski A, Chowell G. Estimating the asymptomatic proportion of coronavirus disease 2019. (COVID-19) cases on board the Diamond Princess cruise ship, Yokohama, Japan, 2020. Eurosurveillance. (2020) 25:1–5. doi: 10.2807/1560-7917.ES.2020.25.10.2000180
PubMed Abstract | CrossRef Full Text
6. Quaresima V, Naldini MM, Cirillo DM. The prospects for the SARS-CoV-2 pandemic in Africa. EMBO Mol Med. (2020) 12:e12488. doi: 10.15252/emmm.202012488
PubMed Abstract | CrossRef Full Text | Google Scholar
7. Ghosh R, Chatterjee S, Dubey S, Lavie CJ. Famotidine against SARS-CoV2: a hope or hype? Mayo Clin Proc. (2020) 95:1797–9. doi: 10.1016/j.mayocp.2020.05.027
CrossRef Full Text | Google Scholar
8. Clerkin KJ, Fried JA, Raikhelkar J, Sayer G, Griffin JM, Masoumi A, et al. COVID-19 and cardiovascular disease. Circulation. (2020) 141:1648–55. doi: 10.1161/CIRCULATIONAHA.120.046941
CrossRef Full Text | Google Scholar
9. Zheng YY, Ma YT, Zhang JY, Xie X. COVID-19 and the cardiovascular system. Nat Rev Cardiol. (2020) 17:259–60. doi: 10.1038/s41569-020-0360-5
PubMed Abstract | CrossRef Full Text | Google Scholar
10. Li G, He X, Zhang L, Ran Q, Wang J, Xiong A, et al. Assessing ACE2 expression patterns in lung tissues in the pathogenesis of COVID-19. J Autoimmun. (2020) 112:102463. doi: 10.1016/j.jaut.2020.102463
PubMed Abstract | CrossRef Full Text | Google Scholar
11. Oudit GY, Crackower MA, Backx PH, Penninger JM. The role of ACE2 in cardiovascular physiology. Trends Cardiovasc Med. (2003) 13:93–101. doi: 10.1016/S1050-1738(02)00233-5
PubMed Abstract | CrossRef Full Text | Google Scholar
12. South AM, Diz DCM. COVID-19, ACE2, and the cardiovascular consequences. Am J Physiol Circ Physiol. (2020) 318:H1084–90. doi: 10.1152/ajpheart.00217.2020
PubMed Abstract | CrossRef Full Text | Google Scholar
13. Klok FA, Kruip MJHA, van der Meer NJM, Arbous MS, Gommers D, Kant KM, et al. Confirmation of the high cumulative incidence of thrombotic complications in critically ill ICU patients with COVID-19: an updated analysis. Thromb Res. (2020) 191:148–50. doi: 10.1016/j.thromres.2020.04.041
PubMed Abstract | CrossRef Full Text | Google Scholar
14. Mehra MR, Desai SS, Kuy SR, Henry TD, Patel AN. Cardiovascular disease, drug therapy, and mortality in COVID-19. N Engl J Med. (2020) 382:E102. doi: 10.1056/NEJMoa2007621
CrossRef Full Text | Google Scholar
15. Mason RJ. Pathogenesis of COVID-19 from a cell biology perspective. Eur Respir J. (2020) 55:9–11. doi: 10.1183/13993003.00607-2020
PubMed Abstract | CrossRef Full Text | Google Scholar
16. Rothan HA, Byrareddy SN. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J Autoimmun. (2020) 109:102433. doi: 10.1016/j.jaut.2020.102433
PubMed Abstract | CrossRef Full Text | Google Scholar
17. Zhang Y, Xiao M, Zhang S, Xia P, Cao W, Jiang W, et al. Coagulopathy and antiphospholipid antibodies in patients with Covid-19. N Engl J Med. (2020) 382:e38. doi: 10.1056/NEJMc2007575
PubMed Abstract | CrossRef Full Text | Google Scholar
18. Cao X. COVID-19: immunopathology and its implications for therapy. Nat Rev Immunol. (2020) 20: 269–70. doi: 10.1038/s41577-020-0308-3
PubMed Abstract | CrossRef Full Text | Google Scholar
19. Duan K, Liu B, Li C, Zhang H, Yu T, Qu J, et al. Effectiveness of convalescent plasma therapy in severe COVID-19 patients. Proc Natl Acad Sci USA. (2020) 117:9490–6. doi: 10.1073/pnas.2007408117
PubMed Abstract | CrossRef Full Text | Google Scholar
20. Cortegiani A, Ingoglia G, Ippolito M, Giarratano A, Einav S. A systematic review on the efficacy and safety of chloroquine for the treatment of COVID-19. J Crit Care. (2020) 57:279–83. doi: 10.1016/j.jcrc.2020.03.005
PubMed Abstract | CrossRef Full Text | Google Scholar
21. Touret F, de Lamballerie X. Of chloroquine and COVID-19. Antiviral Res. (2020) 177:104762. doi: 10.1016/j.antiviral.2020.104762
PubMed Abstract | CrossRef Full Text | Google Scholar
22. Sun D. Remdesivir for treatment of COVID-19: combination of pulmonary and IV administration may offer aditional benefit. AAPS J. (2020) 22:77. doi: 10.1208/s12248-020-00459-8
PubMed Abstract | CrossRef Full Text | Google Scholar
23. de Wit E, Feldmann F, Cronin J, Jordan R, Okumura A, Thomas T, et al. Prophylactic and therapeutic remdesivir (GS-5734) treatment in the rhesus macaque model of MERS-CoV infection. Proc Natl Acad Sci USA. (2020) 117:6771–6. doi: 10.1073/pnas.1922083117
PubMed Abstract | CrossRef Full Text | Google Scholar
24. Wang Y, Zhang D, Du G, Du R, Zhao J, Jin Y, et al. Remdesivir in adults with severe COVID-19: a randomised, double-blind, placebo-controlled, multicentre trial. Lancet. (2020) 395:1569–78. doi: 10.1016/S0140-6736(20)31022-9
PubMed Abstract | CrossRef Full Text | Google Scholar
25. Kang CK, Seong MW, Choi SJ, Kim TS, Choe PG, Song SH, et al. In vitro activity of lopinavir/ritonavir and hydroxychloroquine against severe acute respiratory syndrome coronavirus 2 at concentrations achievable by usual doses. Korean J Intern Med. (2020) 35:782–7. doi: 10.3904/kjim.2020.157
PubMed Abstract | CrossRef Full Text | Google Scholar
26. Rawizza HE, Darin KM, Oladokun R, Brown B, Ogunbosi B, David N, et al. Safety and efficacy of rifabutin among HIV/TB-coinfected children on lopinavir/ritonavir-based ART. J Antimicrob Chemother. (2019) 74:2707–15. doi: 10.1093/jac/dkz219
PubMed Abstract | CrossRef Full Text | Google Scholar
27. Mozzini C, Girelli D. The role of neutrophil extracellular traps in Covid-19: only an hypothesis or a potential new field of research? Thromb Res. (2020) 191:26–7. doi: 10.1016/j.thromres.2020.04.031
CrossRef Full Text | Google Scholar
28. Dunbar JP, Sulpice R, Dugon MM. The kiss of (cell) death: can venom-induced immune response contribute to dermal necrosis following arthropod envenomations? Clin Toxicol. (2019) 57:677–85. doi: 10.1080/15563650.2019.1578367
PubMed Abstract | CrossRef Full Text | Google Scholar
29. Péterfi O, Boda F, Szabó Z, Ferencz E, Bába L. Hypotensive snake venom components-a mini-Review. Molecules. (2019) 24:1–16. doi: 10.3390/molecules24152778
PubMed Abstract | CrossRef Full Text | Google Scholar
30. Hermann K, Ring J. The renin angiotensin system and hymenoptera venom anaphylaxis. Clin Exp Allergy. (1993) 23:762–9. doi: 10.1111/j.1365-2222.1993.tb00364.x
PubMed Abstract | CrossRef Full Text | Google Scholar
31. Sun Y, Han M, Shen Z, Huang H, Miao X. Anti-hypertensive and cardioprotective effects of a novel apitherapy formulation via upregulation of peroxisome proliferator-activated receptor-α and -γ in spontaneous hypertensive rats. Saudi J Biol Sci. (2018) 25:213–9. doi: 10.1016/j.sjbs.2017.10.010
PubMed Abstract | CrossRef Full Text | Google Scholar
32. Kim CMH. Apitherapy – bee venom therapy. In: Biotherapy – History, Principles and Practice. Heidelberg: Springer (2013). p. 77–112. doi: 10.1007/978-94-007-6585-6_4
CrossRef Full Text | Google Scholar
33. Hauser RA, Daguio M, Wester D, Hauser M, Kirchman A, Skinkis C. Bee-venom therapy for treating multiple sclerosis: a clinical trial. Altern Complement Ther. (2001) 7:37–45. doi: 10.1089/107628001300000714
CrossRef Full Text | Google Scholar
34. Alvarez-Fischer D, Noelker C, Vulinović F, Grünewald A, Chevarin C, Klein C, et al. Bee venom and its component apamin as neuroprotective agents in a parkinson disease mouse model. PLoS ONE. (2013) 84:e61700. doi: 10.1371/journal.pone.0061700
PubMed Abstract | CrossRef Full Text | Google Scholar
35. Beck BF. Bee venom therapy. Bee Venom Therapy. Graphic Publishing Company. (1981). p. 238. Available online at: https://www.cabdirect.org/cabdirect/abstract/19820213710
36. Pfaar O, Klimek L, Jutel M, Akdis C, Bousquet J, Akdis M, et al. Handling of allergen immunotherapy in the COVID-19 pandemic: an ARIA-EAACI statement. Allergy. (2020) 75:1546–54. doi: 10.1111/all.14336
PubMed Abstract | CrossRef Full Text | Google Scholar
37. Block J. High risk COVID-19: potential intervention at multiple points in the COVID-19 disease process via prophylactic treatment with azithromycin or bee derived products. Preprints. (2020) 2020040013. doi: 10.20944/preprints202004.0013.v1
CrossRef Full Text | Google Scholar
38. Muller U, Thurnheer U, Patrizzii R, Spies J, Hoigne R. Immunotherapy in bee sting hypersensitivity: bee venom versus whole body extract. Allergy. (1979) 34:369–78. doi: 10.1111/j.1398-9995.1979.tb02006.x
PubMed Abstract | CrossRef Full Text | Google Scholar
39. Marichal T, Starkl P, Reber LL, Kalesnikoff J, Oettgen HC, Tsai M, et al. A beneficial role for immunoglobulin E in host defense against honeybee venom. Immunity. (2013) 39:963–75. doi: 10.1016/j.immuni.2013.10.005
PubMed Abstract | CrossRef Full Text | Google Scholar
40. Johansen P, Senti G, Martinez Gomez JM, Storni T, Beust BR, Wuthrich B, et al. Toll-like receptor ligands as adjuvants in allergen-specific immunotherapy. Clin Exp Allergy. (2005) 35:1591–8. doi: 10.1111/j.1365-2222.2005.02384.x
PubMed Abstract | CrossRef Full Text | Google Scholar
41. Caramalho I, Melo A, Pedro E, Barbosa MMP, Victorino RMM, Pereira Santos MC, et al. Bee venom enhances the differentiation of human regulatory T cells. Allergy. (2015) 70:1340–5. doi: 10.1111/all.12691
PubMed Abstract | CrossRef Full Text | Google Scholar
42. Kim H, Keum DJ, Kwak JW, Chung H-S, Bae H. Bee venom phospholipase a2 protects against acetaminophen-induced acute liver injury by modulating regulatory T cells and IL-10 in mice. PLoS ONE. (2014) 9:e114726. doi: 10.1371/journal.pone.0114726
PubMed Abstract | CrossRef Full Text | Google Scholar
43. Pereira-Santos MC, Baptista AP, Melo A, Alves RR, Soares RS, Pedro E, et al. Expansion of circulating Foxp3+CD25bright CD4 + T cells during specific venom immunotherapy. Clin Exp Allergy. (2008) 38:291–7. doi: 10.1111/j.1365-2222.2007.02887.x
PubMed Abstract | CrossRef Full Text | Google Scholar
44. Lee G, Bae H. bee venom phospholipase A2: yesterday’s enemy becomes today’s friend. Toxins. (2016) 8:48. doi: 10.3390/toxins8020048
PubMed Abstract | CrossRef Full Text | Google Scholar
45. Yang J, Zheng Y, Gou X, Pu K, Chen Z, Guo Q, et al. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: a systematic review and meta-analysis. Int J Infect Dis. (2020) 94:91–5. doi: 10.1016/j.ijid.2020.03.017
PubMed Abstract | CrossRef Full Text | Google Scholar
46. Taketomi Y, Ueno N, Kojima T, Sato H, Murase R, Yamamoto K, et al. Mast cell maturation is driven via a group III phospholipase A 2-prostaglandin D2-DP1 receptor paracrine axis. Nat Immunol. (2013) 14:554–63. doi: 10.1038/ni.2586
PubMed Abstract | CrossRef Full Text | Google Scholar
47. Lambeau G, Lazdunski M. Receptors for a growing family of secreted phospholipases A2. Trends Pharmacol Sci. (1999) 20:162–70. doi: 10.1016/S0165-6147(99)01300-0
PubMed Abstract | CrossRef Full Text | Google Scholar
48. Park S, Baek H, Jung KH, Lee G, Lee H, Kang GH, et al. Bee venom phospholipase A2 suppresses allergic airway inflammation in an ovalbumin-induced asthma model through the induction of regulatory T cells. Immun Inflamm Dis. (2015) 3:386–97. doi: 10.1002/iid3.76
PubMed Abstract | CrossRef Full Text | Google Scholar
49. Zahirović A, Luzar J, Molek P, Kruljec N, Lunder M. Bee venom immunotherapy: current status and future directions. Clin Rev Allergy Immunol. (2020) 58:326–41. doi: 10.1007/s12016-019-08752-x
PubMed Abstract | CrossRef Full Text | Google Scholar
50. Bellinghausen I, Metz G, EnkA H, Christmann S, Knop J, Saloga J. Insect venom immunotherapy induces interleukin-10 production and a Th2-to-Th1 shift, and changes surface marker expression in venom-allergic subjects. Eur J Immunol. (1997). 27:1131–9. doi: 10.1002/eji.1830270513
PubMed Abstract | CrossRef Full Text | Google Scholar
51. ErŽen R, Košnik M, Šilar M, Korošec P. Basophil response and the induction of a tolerance in venom immunotherapy: along term sting challenge study. Allergy. (2012) 67:822–30. doi: 10.1111/j.1398-9995.2012.02817.x
PubMed Abstract | CrossRef Full Text | Google Scholar
52. Jutel M, Pichler WJ, Skrbic D, Urwyler A, Dahinden C, Müller U. Bee venom immunotherapy results in decrease of IL-4 and IL-5 and increase of IFN-gamma secretion in specific allergen-stimulated Tcell cultures. J Immunol. (1995) 154:4187–94. Available online at: https://www.jimmunol.org/content/154/8/4187.long
PubMed Abstract | Google Scholar
53. Jakob T, Rauber MM, Perez-Riverol A, Spillner E, Blank S. The honeybee venom major allergen Api m 10 (Icarapin) and its role in diagnostics and treatment of hymenoptera venom allergy. Curr Allergy Asthma Rep. (2020) 20:48. doi: 10.1007/s11882-020-00943-3
PubMed Abstract | CrossRef Full Text | Google Scholar
54. Shen L, Lee JH, Joo JC, Park SJ, Song J. Bee venom acupuncture for shoulder pain: a systematic review and meta-analysis of randomized controlled trials. J Pharmacopuncture. (2020) 23:44–53. doi: 10.3831/KPI.2020.23.008
PubMed Abstract | CrossRef Full Text | Google Scholar
55. Bramwell VW, Somavarapu S, Outschoorn I AH. Adjuvant action of melittin following intranasal immunisation with tetanus and diphtheria toxoids. J Drug Target. (2003) 11:525–30. doi: 10.1080/10611860410001670080
PubMed Abstract | CrossRef Full Text | Google Scholar
56. Memariani H, Memariani M, Moravvej H, Shahidi-Dadras M. Melittin: a venom-derived peptide with promising anti-viral properties. Eur J Clin Microbiol Infect Dis. (2020) 39:5–17. doi: 10.1007/s10096-019-03674-0
PubMed Abstract | CrossRef Full Text | Google Scholar
57. Pucca MB, Cerni FA, Oliveira IS, Jenkins TP, Argemí L, Sørensen C V. Bee updated: current knowledge on bee venom and bee envenoming therapy. Front Immunol. (2019) 10:2090. doi: 10.3389/fimmu.2019.02090
PubMed Abstract | CrossRef Full Text | Google Scholar
58. Roy A, Bharadvaja N. Venom-derived bioactive compounds as potential anticancer agents: a review. Int J Pept Res Ther. (2020) doi: 10.1007/s10989-020-10073-z
CrossRef Full Text | Google Scholar
59. An WW, Gong XF, Wang MW, Tashiro S, Onodera S, Ikejima T. Norcantharidin induces apoptosis in HeLa cells through caspase, MAPK and mitochondrial pathways. Acta Pharmacol Sin. (2004) 25:1502–08. Available online at: http://www.chinaphar.com/article/view/8413/9071
PubMed Abstract | Google Scholar
60. Cho H-J, Jeong Y-J, Park K-K, Park Y-Y, Chung I-K, Lee K-G, et al. Bee venom suppresses PMA-mediated MMP-9 gene activation via JNK/p38 and NF-κB-dependent mechanisms. J Ethnopharmacol. (2010) 127:662–8. doi: 10.1016/j.jep.2009.12.007
PubMed Abstract | CrossRef Full Text | Google Scholar
61. Baek YH, Huh JE, Lee JD, Choi DY, Park DS. Antinociceptive effect and the mechanism of bee venom acupuncture (Apipuncture) on inflammatory pain in the rat model of collagen-induced arthritis: mediation by α2-Adrenoceptors. Brain Res. (2006) 1073–1074:305–10. doi: 10.1016/j.brainres.2005.12.086
PubMed Abstract | CrossRef Full Text | Google Scholar
62. Choi J, Jeon C, Lee J, Jang J, Quan F, Lee K, et al. Suppressive effects of bee venom acupuncture on paclitaxel-induced neuropathic pain in rats: mediation by spinal α2-adrenergic receptor. Toxins. (2017) 9:351. doi: 10.3390/toxins9110351
PubMed Abstract | CrossRef Full Text | Google Scholar
63. Chirumbolo S, Zanoni G, Ortolani R, Vella A. In vitro biphasic effect of honey bee venom on basophils from screened healthy blood donors. Allergy Asthma Immunol Res. (2011) 3:58. doi: 10.4168/aair.2011.3.1.58
PubMed Abstract | CrossRef Full Text
64. Gu H, Kim W-H, An H, Kim J, Gwon M, Han SM, et al. Therapeutic effects of bee venom on experimental atopic dermatitis. Mol Med Rep. (2018) 18:3711–8. doi: 10.3892/mmr.2018.9398
PubMed Abstract | CrossRef Full Text | Google Scholar
65. Bostanci N, Belibasakis GN. Porphyromonas gingivalis: an invasive and evasive opportunistic oral pathogen. FEMS Microbiol Lett. (2012) 333:1–9. doi: 10.1111/j.1574-6968.2012.02579.x
PubMed Abstract | CrossRef Full Text | Google Scholar
66. Putz T, Ramoner R, Gander H, Rahm A, Bartsch G, Thurnher M. Antitumor action and immune activation through cooperation of bee venom secretory phospholipase A2 and phosphatidylinositol-(3,4)-bisphosphate. Cancer Immunol Immunother. (2006) 55:1374–83. doi: 10.1007/s00262-006-0143-9
PubMed Abstract | CrossRef Full Text | Google Scholar
67. Son D, Lee J, Lee Y, Song H, Lee C, Hong J. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol Ther. (2007) 115:246–70. doi: 10.1016/j.pharmthera.2007.04.004
PubMed Abstract | CrossRef Full Text | Google Scholar
68. Palm NW, Rosenstein RK, Medzhitov R. Allergic host defences. Nature. (2012) 484:465–72. doi: 10.1038/nature11047
PubMed Abstract | CrossRef Full Text | Google Scholar
69. Jilek S, Barbey C, Spertini F, Corthésy B. Antigen-independent suppression of the allergic immune response to bee venom phospholipase A 2 by DNA vaccination in CBA/J mice. J Immunol. (2001) 166:3612–21. doi: 10.4049/jimmunol.166.5.3612
PubMed Abstract | CrossRef Full Text | Google Scholar
70. Palm NW, Rosenstein RK, Yu S, Schenten DD, Florsheim E, Medzhitov R. Bee venom phospholipase A2 induces a primary type 2 response that is dependent on the receptor ST2 and confers protective immunity. Immunity. (2013) 39:976–85. doi: 10.1016/j.immuni.2013.10.006
PubMed Abstract | CrossRef Full Text | Google Scholar
71. An HJ, Lee WR, Kim KH, Kim JY, Lee SJ, Han SM, et al. Inhibitory effects of bee venom on propionibacterium acnes-induced inflammatory skin disease in an animal model. Int J Mol Med. (2014) 34:1341–8. doi: 10.3892/ijmm.2014.1933
PubMed Abstract | CrossRef Full Text | Google Scholar
72. Rizk JG, Kalantar-Zadeh K, Mehra MR, Lavie CJ, Rizk Y, Forthal DN. Pharmaco-Immunomodulatory therapy in COVID-19. Drugs. (2020) 80:1267–92. doi: 10.1007/s40265-020-01367-z
CrossRef Full Text | Google Scholar
73. Babon A, Almunia C, Boccaccio C, Beaumelle B, Gelb MH, Ménez A, et al. Cross-presentation of a CMV pp65 epitope by human dendritic cells using bee venom PLA 2 as a membrane-binding vector. FEBS Lett. (2005) 579:1658–64. doi: 10.1016/j.febslet.2005.02.019
PubMed Abstract | CrossRef Full Text | Google Scholar
74. Almunia C, Bretaudeau M, Held G, Babon A, Marchetti C, Castelli FA, et al. Bee venom phospholipase A2, a good “Chauffeur” for delivering tumor antigen to the MHC I and MHC II peptide-loading compartments of the dendritic cells: the case of NY-ESO-1. PLoS ONE. (2013) 8:1–17. doi: 10.1371/journal.pone.0067645
PubMed Abstract | CrossRef Full Text | Google Scholar
75. Eltahir Saeed WS, Gasim Khalil EA. Immune response modifying effects of bee venom protein [Melittin]/Autoclaved L. donovani complex in CD1 Mice: the search for new vaccine adjuvants. J Vaccines Vaccin. (2017) 08:6–11. doi: 10.4172/2157-7560.1000372
CrossRef Full Text | Google Scholar
76. Cherniack EP, Govorushko S. To bee or not to bee: the potential efficacy and safety of bee venom acupuncture in humans. Toxicon. (2018) 154:74–8. doi: 10.1016/j.toxicon.2018.09.013
CrossRef Full Text | Google Scholar
77. Seo B-K, Han K, Kwon O, Jo D-J, Lee J-H. Efficacy of bee venom acupuncture for chronic low back pain: a randomized, double-blinded, sham-controlled trial. Toxins. (2017) 9:361. doi: 10.3390/toxins9110361
PubMed Abstract | CrossRef Full Text | Google Scholar
78. Gu H, Han SM, Park K-K. therapeutic effects of apamin as a bee venom component for non-neoplastic disease. Toxins. (2020) 12:195. doi: 10.3390/toxins12030195
PubMed Abstract | CrossRef Full Text | Google Scholar
79. Rady I, Siddiqui IA, Rady M, Mukhtar H. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. Cancer Lett. (2017) 402:16–31. doi: 10.1016/j.canlet.2017.05.010
PubMed Abstract | CrossRef Full Text | Google Scholar
80. Son DJ, Kang J, Kim TJ, Song HS, Sung KJ, Yun DY, et al. Melittin, a major bioactive component of bee venom toxin, inhibits PDGF receptor beta-tyrosine phosphorylation and downstream intracellular signal transduction in rat aortic vascular smooth muscle cells. J Toxicol Environm Health Part A. (2007) 70:1350–5. doi: 10.1080/15287390701428689
PubMed Abstract | CrossRef Full Text | Google Scholar
81. Zhang S, Liu Y, Ye Y, Wang XR, Lin LT, Xiao LY, et al. Bee venom therapy: potential mechanisms and therapeutic applications. Toxicon. (2018) 148:64–73. doi: 10.1016/j.toxicon.2018.04.012
PubMed Abstract | CrossRef Full Text | Google Scholar
82. King TP, Jim SY, Wittkowski K. Inflammatory role of two venom components of yellow jackets (Vespula vulgaris): A mast cell degranulating peptide mastoparan and phospholipase A1. Int Arch Allergy Immunol. (2003) 131:25–32. doi: 10.1159/000070431
PubMed Abstract | CrossRef Full Text | Google Scholar
83. LaFerla FM, Green KN, Oddo S. Intracellular amyloid-beta in alzheimer’s disease. Nat Rev Neurosci. (2007) 8:499–509. doi: 10.1038/nrn2168
PubMed Abstract | CrossRef Full Text | Google Scholar
84. Shkenderov S, Koburova K. Adolapin-A newly isolated analgetic and anti-inflammatory polypeptide from bee venom. Toxicon. (1982) 20:317–21. doi: 10.1016/0041-0101(82)90234-3
PubMed Abstract | CrossRef Full Text | Google Scholar
85. Moreno M, Giralt E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: melittin, apamin and mastoparan. Toxins. (2015) 7:1126–50. doi: 10.3390/toxins7041126
PubMed Abstract | CrossRef Full Text | Google Scholar
86. Bilò MB, Bonifazi F. The natural history and epidemiology of insect venom allergy: clinical implications. Clin Exp Allergy. (2009) 39:1467–76. doi: 10.1111/j.1365-2222.2009.03324.x
PubMed Abstract | CrossRef Full Text | Google Scholar
87. Antonicelli L, Bilò MB, Bonifazi F. Epidemiology of hymenoptera allergy. Curr Opin Allergy Clin Immunol. (2002) 2:341–6. doi: 10.1097/00130832-200208000-00008
PubMed Abstract | CrossRef Full Text | Google Scholar
88. Mingomataj EÇ, Bakiri AH. Episodic hemorrhage during honeybee venom anaphylaxis: potential mechanisms. J Investig Allergol Clin Immunol. (2012) 22:237–44. Available online at: http://www.jiaci.org/issues/vol22issue4/vol22issue04-1.html
PubMed Abstract | Google Scholar
89. White J. Venomous Animals: Clinical Toxinology. In: EXS. (2010) p. 233–91. Available online at: http://www.ncbi.nlm.nih.gov/pubmed/20358686 doi: 10.1007/978-3-7643-8338-1_7
CrossRef Full Text | Google Scholar
90. Vetter RS, Visscher PK. Bites and stings of medically important venomous arthropods. Int J Dermatol. (1998) 37:481–96. doi: 10.1046/j.1365-4362.1998.00455.x
PubMed Abstract | CrossRef Full Text | Google Scholar
91. Lima PR, de Brochetto-Braga MR. Hymenoptera venom review focusing on Apis mellifera. J Venom Anim Toxins Incl Trop Dis. (2003) 9:149–62. doi: 10.1590/S1678-91992003000200002
CrossRef Full Text | Google Scholar
92. Golden DBK. Epidemiology of allergy to insect venoms and stings. Allergy Asthma Proc. (1989) 10:103–7. doi: 10.2500/108854189778960964
PubMed Abstract | CrossRef Full Text | Google Scholar
93. Stoevesandt J, Sturm GJ, Bonadonna P, Oude Elberink JNG, Trautmann A. Risk factors and indicators of severe systemic insect sting reactions. Allergy Eur J Allergy Clin Immunol. (2020) 75:535–45. doi: 10.1111/all.13945
PubMed Abstract | CrossRef Full Text | Google Scholar
94. Reber LL, Hernandez JD, Galli SJ. The pathophysiology of anaphylaxis. J Allergy Clin Immunol. (2017) 140:335–48. doi: 10.1016/j.jaci.2017.06.003
PubMed Abstract | CrossRef Full Text | Google Scholar
95. Chen J, Guan S-M, Sun W, Fu H. Melittin, the major pain-producing substance of bee venom. Neurosci Bull. (2016) 32:265–72. doi: 10.1007/s12264-016-0024-y
PubMed Abstract | CrossRef Full Text | Google Scholar
96. Abd El-Wahed AA, Khalifa SAM, Sheikh BY, Farag MA, Saeed A, Larik FA, et al. Bee Venom Composition: From Chemistry to Biological Activity. Stud Nat Prod Chem. (2019) 60:459–84. doi: 10.1016/B978-0-444-64181-6.00013-9
CrossRef Full Text | Google Scholar
97. Stone KD, Prussin C, Metcalfe DD. IgE, mast cells, basophils, and eosinophils. J Allergy Clin Immunol. (2010) 125:S73–80. doi: 10.1016/j.jaci.2009.11.017
PubMed Abstract | CrossRef Full Text | Google Scholar
98. Schumacher M, Tveten M, Egen N. Rate and quantity of delivery of venom from honeybee stings. J Allergy Clin Immunol. (1994). 93:831–5. doi: 10.1016/0091-6749(94)90373-5
PubMed Abstract | CrossRef Full Text | Google Scholar
99. Toledo LFM, Moore DCBC, Caixeta DMDL, Salú MDS, Farias CVB, Azevedo ZMA. Multiple bee stings, multiple organs involved: a case report. Rev Soc Bras Med Trop. (2018). 51:560–2. doi: 10.1590/0037-8682-0341-2017
PubMed Abstract | CrossRef Full Text | Google Scholar
100. Rajendiran C, Puvanalingam A, Thangam D, Ragunanthanan S, Ramesh D, Venkatesan S, et al. Stroke after multiple bee sting. J Assoc Physicians India. (2012) 60:122–4.
PubMed Abstract | Google Scholar
101. Bhalotia S, Kumar NR, Kaur J, Devi A. Honey bee venom and its composition: focusing on different apis species-a review. J Basic Appl Eng Res. (2016) 3:96–8. Available online at: https://www.krishisanskriti.org/vol_image/10Jun2016100659z47%20%20%20%20%20%20%20%20%20%20%20Anita%20Devi%202%20%20%20%20%20%20%20%20%20%20%2096-98.pdf
102. Hossen MS, Gan SH, Khalil MI. Melittin, a potential natural toxin of crude bee venom: probable future arsenal in the treatment of diabetes mellitus. J Chem. (2017) 2017:1–7. doi: 10.1155/2017/4035626
CrossRef Full Text | Google Scholar
103. Wehbe R, Frangieh J, Rima M, El Obeid D, Sabatier J-M, Fajloun Z. Bee venom: overview of main compounds and bioactivities for therapeutic interests. Molecules. (2019) 24:2997. doi: 10.3390/molecules24162997
PubMed Abstract | CrossRef Full Text | Google Scholar
104. Mokosuli YS, Repi RA, Worang RL, Mokosuli C, Semuel Y. Potential antioxidant and anticancer effect of apis dorsata binghami crude venom from minahasa, north sulawesi. J Entomol Zool Stud JEZS. (2017) 112:112–9. Available online at: https://www.entomoljournal.com/archives/?year=2017&vol=5&issue=2&ArticleId=1581
105. Bachmayer H, Kreil G, Suchanek G. Synthesis of promelittin and melittin in the venom gland of queen and worker bees: patterns observed during maturation. J Insect Physiol. (1972) 18:1515–21. doi: 10.1016/0022-1910(72)90230-2
CrossRef Full Text | Google Scholar
106. Abusabbah M, Hong Lau W, Mahmoud ME, Salih AM, Omar D. Prospects of using carbohydrates as supplemented-diets and protein rich mixture as alternative-diet to improve the quality of venom produced by Apis cerana L. J Entomol Zool Stud. (2016) 4:23–6.
107. Abdela N, Jilo K. Bee venom and its therapeutic values: a review. Adv Life Sci Technol. (2016) 44:18–22. Available online at: https://www.iiste.org/Journals/index.php/ALST/article/view/30404/31249
108. Owen MD, Pfaff LA. Melittin synthesis in the venom system of the honey bee (Apis mellifera L). Toxicon. (1995) 33:1181–8. doi: 10.1016/0041-0101(95)00054-P
PubMed Abstract | CrossRef Full Text | Google Scholar
109. Leandro LF, Mendes CA, Casemiro LA, Vinholis AHC, Cunha WR, Almeida R de, et al. Antimicrobial activity of apitoxin, melittin and phospholipase A2 of honey bee (Apis mellifera) venom against oral pathogens. An Acad Bras Cienc. (2015) 87:147–55. doi: 10.1590/0001-3765201520130511
PubMed Abstract | CrossRef Full Text | Google Scholar
110. Uddin MB, Lee B-H, Nikapitiya C, Kim J-H, Kim T-H, Lee H-C, et al. Inhibitory effects of bee venom and its components against viruses in vitro and in vivo. J Microbiol. (2016) 54:853–66. doi: 10.1007/s12275-016-6376-1
PubMed Abstract | CrossRef Full Text | Google Scholar
111. Jung GB, Huh J-E, Lee H-J, Kim D, Lee G-J, Park H-K, et al. Anti-cancer effect of bee venom on human MDA-MB-231 breast cancer cells using Raman spectroscopy. Biomed Opt Express. (2018) 9:5703 doi: 10.1364/BOE.9.005703
PubMed Abstract | CrossRef Full Text | Google Scholar
112. Sabaratnam V, Gurunathan S, Raman J, Abd Malek SN, John P. Green synthesis of silver nanoparticles using Ganoderma neo-japonicum Imazeki: a potential cytotoxic agent against breast cancer cells. Int J Nanomedicine. (2013) 8:4399. doi: 10.2147/IJN.S51881
CrossRef Full Text | Google Scholar
113. Hong J, Lu X, Deng Z, Xiao S, Yuan B, Yang K. How melittin inserts into cell membrane: conformational changes, inter-peptide cooperation, and disturbance on the membrane. Molecules. (2019) 24:1775. doi: 10.3390/molecules24091775
PubMed Abstract | CrossRef Full Text | Google Scholar
114. Liu S, Yu M, He Y, Xiao L, Wang F, Song C, et al. Melittin prevents liver cancer cell metastasis through inhibition of the Rac1-dependent pathway. Hepatology. (2008) 47:1964–73. doi: 10.1002/hep.22240
PubMed Abstract | CrossRef Full Text | Google Scholar
115. Gupta T, Gupta SK. Potential adjuvants for the development of a SARS-CoV-2 vaccine based on experimental results from similar coronaviruses. Int Immunopharmacol. (2020) 86:106717. doi: 10.1016/j.intimp.2020.106717
PubMed Abstract | CrossRef Full Text | Google Scholar
116. Okba NM, Raj VS, Haagmans BL. Middle East respiratory syndrome coronavirus vaccines: current status and novel approaches. Curr Opin Virol. (2017) 23:49–58. doi: 10.1016/j.coviro.2017.03.007
PubMed Abstract | CrossRef Full Text | Google Scholar
117. Vijay R, Hua X, Meyerholz DK, Miki Y, Yamamoto K, Gelb M, et al. Critical role of phospholipase A2 group IID in age-related susceptibility to severe acute respiratory syndrome-CoV infection. J Exp Med. (2015) 212:1851–68. doi: 10.1084/jem.20150632
PubMed Abstract | CrossRef Full Text | Google Scholar
118. Mousavizadeh L, Ghasemi S. Genotype and phenotype of COVID-19: Their roles in pathogenesis. J Microbiol Immunol Infect. (2020) 2020:5. doi: 10.1016/j.jmii.2020.03.022
PubMed Abstract | CrossRef Full Text | Google Scholar
119. Dawood AA. Mutated COVID-19 may foretell a great risk for mankind in the future. New Microbes New Infect. (2020) 35:100673. doi: 10.1016/j.nmni.2020.100673
PubMed Abstract | CrossRef Full Text
120. de Haan CAM, Kuo L, Masters PS, Vennema H, Rottier PJM. Coronavirus particle assembly: primary structure requirements of the membrane protein. J Virol. (1998) 72:6838–50. doi: 10.1128/JVI.72.8.6838-6850.1998
PubMed Abstract | CrossRef Full Text | Google Scholar
121. Fenard D, Lambeau G, Valentin E, Lefebvre J-C, Lazdunski M, Doglio A. Secreted phospholipases A2, a new class of HIV inhibitors that block virus entry into host cells. J Clin Invest. (1999) 104:611–8. doi: 10.1172/JCI6915
PubMed Abstract | CrossRef Full Text | Google Scholar
122. Soman NR, Baldwin SL, Hu G, Marsh JN, Lanza GM, Heuser JE, et al. Molecularly targeted nanocarriers deliver the cytolytic peptide melittin specifically to tumor cells in mice, reducing tumor growth. J Clin Invest. (2009) 119:2830–42. doi: 10.1172/JCI38842
PubMed Abstract | CrossRef Full Text | Google Scholar
123. Youngren-Ortiz SR, Chougule MB, Morris KR. Development and evaluation of siRNA loaded gelatin nanocarriers for the treatment of asthma. Dissertations and Theses. University of Hawaii at Hilo. (2016) Available online at: https://dspace.lib.hawaii.edu/handle/10790/2758
124. Yang W, Hu F, Xu X. Bee venom and SARS-CoV-2. Toxicon. (2020) 181:69–70. doi: 10.1016/j.toxicon.2020.04.105
PubMed Abstract | CrossRef Full Text | Google Scholar
125. Balozet L, Bücherl W, Klobusitzky D De, Valle JR, Halstead BW, Mcmichael DF. Contributors to this volume venomous animals. (1971) 3:1–459.
126. Casewell NR, Wüster W, Vonk FJ, Harrison RA, Fry BG. Complex cocktails: the evolutionary novelty of venoms. Trends Ecol Evol. (2013) 28:219–29. doi: 10.1016/j.tree.2012.10.020
PubMed Abstract | CrossRef Full Text | Google Scholar
127. Garraud O, Hozzein WN, Badr G. Wound healing: time to look for intelligent, ‘natural’ immunological approaches? BMC Immunol. (2017) 18:23. doi: 10.1186/s12865-017-0207-y
PubMed Abstract | CrossRef Full Text | Google Scholar
128. Ali M. Studies on bee venom and its medical uses. Int J Adv Res. (2012) 1:1–15. Available online at: http://www.ijoart.org/docs/Studies-on-Bee-Venom-and-Its-Medical-Uses.pdf
129. Seif F, Khoshmirsafa M, Aazami H, Mohsenzadegan M, Sedighi G, Bahar M. The role of JAK-STAT signaling pathway and its regulators in the fate of T helper cells. Cell Commun Signal. (2017) 15:23. doi: 10.1186/s12964-017-0177-y
PubMed Abstract | CrossRef Full Text | Google Scholar
130. Dams D, Briers Y. Enzybiotics: enzyme-based antibacterials as therapeutics. Adv Exp Med Biol. (2019) 1148:233–53. doi: 10.1007/978-981-13-7709-9_11
PubMed Abstract | CrossRef Full Text | Google Scholar
131. Boens S, Szekér K, Eynde A Van, Bollen M. Phosphatase Modulators. In: Millán JL, editor. Methods in Molecular Biology. Totowa, NJ: Humana Press. (2013). p.271–81. doi: 10.1007/978-1-62703-562-0_16
CrossRef Full Text | Google Scholar
132. Szulc P, Bauer DC. Biochemical markers of bone turnover in osteoporosis. In: Osteoporosis. Elsevier (2013). p. 1573–610. doi: 10.1016/B978-0-12-415853-5.00067-4. Available online at: https://www.sciencedirect.com/science/article/pii/B9780124158535000674?via%3Dihub
CrossRef Full Text | Google Scholar
133. Murakami M, Nakatani Y, Atsumi G, Inoue K, Kudo I. Regulatory functions of phospholipase A2. Crit Rev Immunol. (2017) 37:121–79. doi: 10.1615/CritRevImmunol.v37.i2-6.20
PubMed Abstract | CrossRef Full Text | Google Scholar
134. Stahelin R V. Chapter 8 – Phospholipid Catabolism. In: Ridgway ND, McLeod L, editors. Lipoproteins and Membranes (Sixth Edition) RSBT-B. Boston: Elsevier. (2016) p. 237–57. doi: 10.1016/B978-0-444-63438-2.00008-0
135. Connolly TM, Lawing WJ, Majerus PW. Protein kinase C phosphorylates human platelet inositol trisphosphate 5′-phosphomonoesterase, increasing the phosphatase activity. Cell. (1986) 46:951–8. doi: 10.1016/0092-8674(86)90077-2
PubMed Abstract | CrossRef Full Text | Google Scholar
136. Brás NF, Santos-Martins D, Fernandes PA, Ramos MJ. Mechanistic pathway on human α-glucosidase maltase-glucoamylase Unveiled by QM/MM calculations. J Phys Chem B. (2018) 122:3889–99. doi: 10.1021/acs.jpcb.8b01321
CrossRef Full Text | Google Scholar
137. Holtsberg FW, Ozgur LE, Garsetti DE, Myers J, Egan RW, Clark MA. Presence in human eosinophils of a lysophospholipase similar to that found in the pancreas. Biochem J. (1995) 309:141–4. doi: 10.1042/bj3090141
PubMed Abstract | CrossRef Full Text | Google Scholar
138. Karamitros CS, Konrad M. Human 60-kDa lysophospholipase contains an N-terminal l-Asparaginase domain that is allosterically regulated by l-Asparagine. J Biol Chem. (2014) 289:12962–75. doi: 10.1074/jbc.M113.545038
PubMed Abstract | CrossRef Full Text | Google Scholar
139. Soliman C, Eastwood S, Truong VK, Ramsland PA, Elbourne A. The membrane effects of melittin on gastric and colorectal cancer. PLoS ONE. (2019) 14:e0224028. doi: 10.1371/journal.pone.0224028
PubMed Abstract | CrossRef Full Text | Google Scholar
140. Issa MF, Tuboly G, Kozmann G, Juhasz Z. Automatic ECG artefact removal from EEG signals. Meas Sci Rev. (2019) 19:101–8. doi: 10.2478/msr-2019-0016
CrossRef Full Text | Google Scholar
141. Elieh Ali Komi D, Shafaghat F, Zwiener RD. Immunology of bee venom. Clin Rev Allergy Immunol. (2018) 54:386–96. doi: 10.1007/s12016-017-8597-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Zdroj: https://www.frontiersin.org/articles/10.3389/fpubh.2020.594458/full#F1