

1. Skupina pre imunomoduláciu a toleranciu, Alergia a klinická imunológia Zápal, oprava a vývoj, Národný inštitút srdca a pľúc, Imperial College London, Londýn, Spojené kráľovstvo
2. Pediatrické oddelenie pre alergiu, Hacettepe University School of Medicine, Ankara, Turecko
3. MRC and Asthma UK Center in Alergic Mechanisms of Asthma, Londýn, Spojené kráľovstvo
Uštipnutie blanokrídlym hmyzom môže spôsobiť lokálne aj systémové alergické reakcie a dokonca aj život ohrozujúcu anafylaxiu. Spolu s farmaceutickými liekmi a potravinami je jed blanokrídlovcov jednou z najčastejších príčin anafylaxie u ľudí. Doteraz nebol identifikovaný žiadny parameter, ktorý by mohol predpovedať, ktorí senzibilizovaní ľudia budú mať v budúcnosti systémovú reakciu na bodnutie (SSR), sú však známe niektoré rizikové faktory, ako je mastocytóza a vek > 40 rokov. Imunoterapia jedom (VIT) je najúčinnejšou metódou liečby ľudí s SSR, ktorá sa ukazuje ako účinná aj po prerušení liečby. Rozvoj periférnej tolerancie je hlavným mechanizmom počas imunoterapie. Je sprostredkovaná produkciou blokujúcich protilátok IgG/IgG4, ktoré môžu inhibovať reakcie závislé od IgE prostredníctvom receptorov IgE s vysokou afinitou (FcεRI) aj s nízkou afinitou (FcεRII) na žírnych bunkách, bazofiloch a B bunkách. Generovanie antigénovo špecifických regulačných T buniek produkuje IL-10 a potláča Th2 imunitu a imunitné reakcie sa posúvajú smerom k reakcii typu Th1. B regulačné bunky sa tiež podieľajú na produkcii IL-10 a rozvoji dlhodobej imunitnej tolerancie. Počas VIT tiež klesá počet efektorových buniek v cieľových orgánoch, ako sú žírne bunky, bazofily, vrodené lymfocyty 2. typu a eozinofily. Viaceré metaanalýzy a randomizované kontrolované štúdie preukázali, že VIT je účinná pri prevencii SSR pred bodnutím a zlepšuje kvalitu života. V tomto prehľade sa diskutuje o riziku SSR pri alergii na jed a ako VIT zmenila toto riziko.
Úvod
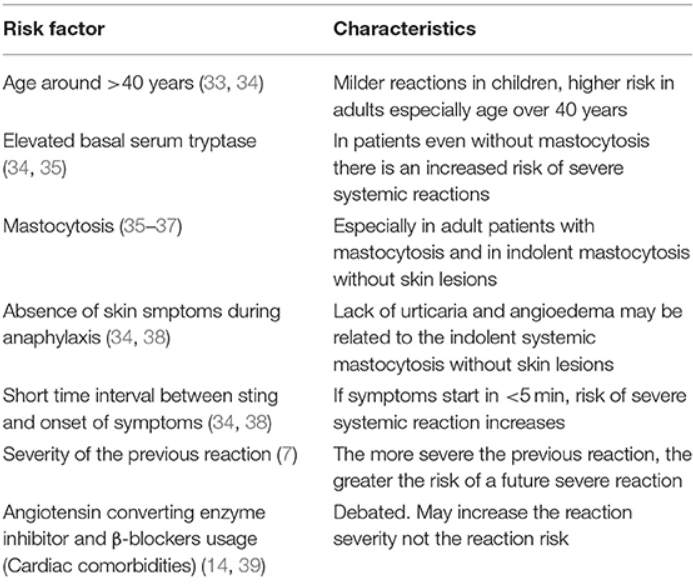
Skupina hmyzu Hymenoptera zahŕňa podskupiny Apidae a Vespidae a tiež Formicidae, čo je nad rámec tohto prehľadu. Apidae pozostáva z druhov Apis mellifera (včely medonosné) a čmeliakov (čmeliakov) a podtrieda Vespidae zahŕňa druhy Vespula („žlté kabátiky“, osy a sršne) a druhy Polistes („papierové“ osy) (1, 2). Uštipnutie včelou nie je vo všeobecnosti silnejšie, ale vstrekuje viac jedu. Včely vstreknú 50 až 140 mikrogramov jedu, zatiaľ čo osy dodajú takmer 3 mikrogramy jedu s každým bodnutím. Včely môžu bodnúť raz, ale osy majú schopnosť bodnúť viackrát (1–3). Alergia na bodnutie hmyzom môže spôsobiť lokálne, veľké lokálne (>10 cm v priemere) alebo dokonca systémové reakcie (SR) a potenciálne život ohrozujúce anafylaktické reakcie (4–6). Miera systémových reakcií na bodnutie v epidemiologických štúdiách v Európe sa pohybovala medzi 0,3 a 7,5 % u dospelých (7) a 0,15 – 3,4 % u detí (7, 8). Pravdepodobnosť SR a možnosť život ohrozujúcej anafylaxie súvisí s mnohými faktormi, vrátane závažnosti predchádzajúcej reakcie, alergie na včelí jed, hladiny základnej sérovej tryptázy a prítomnosti mastocytózy, zvýšenej aktivácie bazofilov, veku a základného zdravotného stavu. podmienky (7).
Imunoterapia jedom (VIT) vedie ku kompletnej ochrane pred SSR v 77–84 % prípadov v prípade včely medonosnej a v 91–96 % v prípade jedu vesieho (9–11). Frekvencia systémových nežiaducich účinkov počas VIT sa pohybuje medzi 8 a 20 % z veľkých multicentrických štúdií (12, 13). V nedávnej štúdii Stoevesandta a kol. miera systémových reakcií 11,7 % (akékoľvek reakcie vrátane subjektívnych) bola hlásená počas fázy budovania VIT; miera SSR však klesla na 3 %, keď sa použili objektívne diagnostické kritériá anafylaxie (14). Najdôležitejšími rizikovými faktormi súvisiacimi so systémovými reakciami počas VIT sú imunoterapia včelím jedom, rýchle zvýšenie dávky počas fázy budovania a pravdepodobne vysoké bazálne hladiny tryptázy pri alergii na veslopie, ale nie pri alergii na včelí jed (9). Ochranný účinok VIT pretrváva roky po ukončení liečby. Dlhodobý výsledok systémových reakcií po prerušení VIT je lepší u detí v porovnaní s dospelými a pre jed veschy v porovnaní s VIT včely (15–17).
Cieľom tohto prehľadu je najprv diskutovať o epidemiológii a rizikových faktoroch anafylaxie hmyzím jedom, potom sa zameriava na mechanizmy VIT na prevenciu SSR po bodnutí hmyzom a nakoniec sa zameriava na diskusiu o účinnosti, bezpečnosti a dlhodobých účinkoch VIT, ako aj o rizikových faktoroch. súvisiace s SSR počas a po VIT.
Epidemiológia alergie na jed a alergických reakcií
Prevalencia bodnutia druhmi Hymenoptera počas života sa pohybuje od 56,6 do 94,5 % u dospelých a 37,5 % u detí do 14 rokov (7, 8). Miera senzibilizácie, indikovaná buď pozitívnym kožným prick testom alebo špecifickou IgE pozitivitou, sa u dospelých pohybuje medzi 9,3 a 28,7 %. V jednej štúdii sa zistilo, že deti sú na 3,7 % pozitívne na druhy blanokrídlovcov (väčšinou včely medonosné) testovaním vpichom do kože (18).
Miera systémových reakcií na bodnutie v epidemiologických štúdiách v Európe sa pohybuje medzi 0,3 a 7,5 % u dospelých (7). Medzi týmito reakciami je frekvencia anafylaktického šoku medzi 0,6 a 42,8 % (18–23). Podľa nedávneho pozičného dokumentu u dospelých sa respiračné a kardiovaskulárne symptómy môžu vyskytnúť až v 70 % systémových reakcií (24). Tento široký rozsah odzrkadľuje nedostatočný konsenzus o definícii anafylaxie, rozdiely v technikách zberu údajov a variabilitu stupňa vystavenia bodnutiu v rôznych klimatických podmienkach (7). U detí je prevalencia SR oveľa nižšia a pohybuje sa od 0,15 do 3,4 % (7, 8). Okrem toho, SR u detí je približne 60 % miernych a obmedzených len na kožu (24).
Včelári sú zraniteľnou a vysoko rizikovou populáciou pre rozvoj alergických reakcií na uštipnutie včelou. V tejto špecifickej skupine sú miery SR vyššie ako v bežnej populácii a pohybujú sa od 14 do 38 % (25). Prijatie viac ako 200 žihadiel za rok takmer úplne chráni pred SR, zatiaľ čo prijatie menej ako 25 žihadiel za rok súvisí s mierou SR 45 % (25). V prieskume britských včelárov sa zistilo, že rizikové faktory pre SR sú ženské pohlavie, pozitívna rodinná anamnéza alergie na včelí jed, premedikácia antihistaminikami pred návštevou úľa a 2 a viac rokov včelárenia pred prvou SR (26).
Podľa Európskej siete závažných alergických reakcií (NORA) bolo 20,2 % všetkých prípadov anafylaxie u detí a 48,2 % dokumentovanej anafylaxie u dospelých spôsobených hmyzím jedom. V tejto štúdii 59 terciárnych centier pre alergiu, dermatológiu a pediatriu z 10 rôznych krajín hlásilo 3 333 prípadov anafylaxie (27). Len 27,6 % prípadov anafylaxie hmyzu dostalo adrenalín na mieste (27). V populačných štúdiách uskutočnených počas prvej dekády 21. storočia sa miera anafylaxie v dôsledku hmyzieho jedu pohybovala od 7,3 do 59 % a bola zistená väčšinou u dospelých (7).
Úmrtia na bodnutie hmyzom boli predtým skúmané v mnohých štúdiách. Štúdia z Kostariky uvádza 52 úmrtí za 22 rokov, čo predstavuje 0,74 úmrtí na milión obyvateľov za rok (28), čo je oveľa viac ako štúdia z USA s počtom 0,14 úmrtí na milión obyvateľov za rok (29). . Nedávna správa zo Spojeného kráľovstva uvádzala 93 úmrtí na anafylaxiu jedom medzi rokmi 1992 a 2012, čo predstavuje 0,09 úmrtí na milión obyvateľov za rok (30). Najmä predchádzajúca anamnéza alergie na blanokrídlovce, mužské pohlavie, vyšší vek a oneskorené podávanie adrenalínu sú dôležitými rizikovými faktormi pre fatálne reakcie (31, 32).
Rizikové faktory súvisiace so SSR uštipnutím blanokrídlym hmyzom sú zhrnuté v tabuľke 1.
Alergia na jed, poruchy aktivácie žírnych buniek a anafylaxia
Mastocytóza je klonálna neoplastická porucha žírnych buniek, ktorá je charakterizovaná predovšetkým kožnými alebo systémovými podtypmi a niekedy zriedkavými formami. Mastocytóza zvyčajne zahŕňa somatickú mutáciu KIT D816V, vykazuje aberantnú expresiu CD25 alebo CD2 na žírnych bunkách a je často sprevádzaná zvýšenou bazálnou sérovou tryptázou (BST) (40). U pacientov s kožnou mastocytózou sa agregáty mastocytov nachádzajú iba v koži a táto forma je väčšinou diagnostikovaná u dojčiat a detí (41). Systémová mastocytóza (SM) sa často vyskytuje u dospelých a je rozdelená do štyroch podtypov (indolentná SM, SM spojená s hematologickou poruchou, agresívna SM a sarkóm mastocytov); najbežnejšou z nich je indolentná systémová mastocytóza (ISM). ISM sa môže prejaviť normálnym BST a nedostatkom kožných lézií (42). Prevalencia mastocytózy u pacientov s alergiou na jed blanokrídleho hmyzu môže byť až 7,9 %, čo je výrazne viac ako u bežnej populácie (3–13/100 000 obyvateľov) (43, 44). Podobne bodnutie blanokrídlovcov je najčastejšou príčinou anafylaxie u ľudí s mastocytózou a prevalencia je takmer 30 % (45). U značného počtu pacientov s ISM môže byť anafylaxia jedu blanokrídlovcov (HVA) prvým príznakom ochorenia. Títo pacienti s ISM bez kožných lézií sú väčšinou muži s nižšími hladinami BST a nižšou frekvenciou mutácií KIT (46). Anafylaxia u pacientov s mastocytózou je väčšinou charakterizovaná srdcovými symptómami, ako je hypotenzia a synkopálne epizódy, zatiaľ čo kožné symptómy, ako je urtikária a angioedém, sú zriedkavé. Preto, ak sa u pacienta s VHA vyskytne hypotenzia a synkopa bez kožných symptómov, treba mať podozrenie na mastocytózu (47). Nedávno bol pre prípady HVA vyvinutý systém hodnotenia s názvom REMA na základe pohlavia, klinických symptómov a BST na identifikáciu pacientov s klonálnym systémovým ochorením žírnych buniek. Skóre ≥2 naznačuje, že ISM by sa malo vylúčiť so senzitivitou 92 % a špecificitou 81 % (47).
Presné mechanizmy zvýšeného výskytu HVA pri mastocytóze nie sú dobre pochopené. Možné vysvetlenia tejto koexistencie sú však nasledovné: (1) Zvýšená záťaž žírnych buniek môže viesť k vyššiemu uvoľňovaniu mediátora v prípade bodnutia; (2) Perivaskulárne umiestnenie mastocytov môže spôsobiť priamy prístup mediátorov do systémového obehu, čo zvyšuje závažnosť reakcie; (3) Zisk funkčnej mutácie, KIT D816V, môže zabrániť apoptóze žírnych buniek a zvýšiť proliferáciu a diferenciáciu žírnych buniek, a napokon (4) Fosfolipáza A2, zložka jedu blanokrídlovcov, môže spôsobiť priame uvoľnenie mediátora zo žírnych buniek. (36).
VIT sa javí ako najvhodnejšia možnosť liečby u pacientov s mastocytózou a HVA. VIT je zvyčajne dobre tolerovaná, bezpečná a účinná. Zatiaľ nie je jasné, či BST zvyšuje riziko nežiaducich udalostí počas VIT. Napriek preferencii celoživotnej VIT u pacientov s mastocytózou neexistuje v literatúre dôkaz o celoživotnej liečbe. Pre istotu sa však odporúča, aby sa VIT predĺžila u pacientov so zvýšenou SBT (>11,4 μg/l) alebo overenou mastocytózou, ak bola počiatočná SSR závažná (9).
Mechanizmy imunoterapie jedom
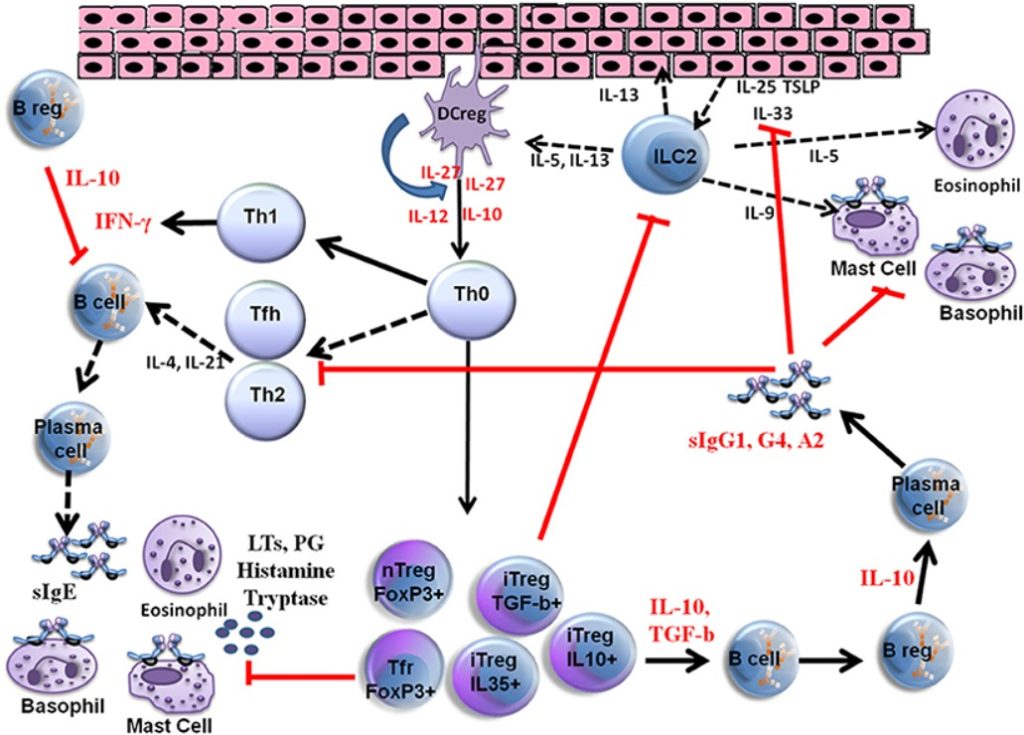
VIT je jediná účinná a imunomodulačná liečba u pacientov s anamnézou anafylaxie a preukázala sa ako účinná u 80 až 95 % pacientov s alergiou na včelí jed a na vespulový jed (9–11). Spočiatku interakcia medzi alergénmi, epitelovými bunkami a dendritickými bunkami spúšťa reakciu produkovanú vrodeným imunitným systémom a následne sa vyvíjajú adaptívne imunitné reakcie. Na rozvoji periférnej imunitnej tolerancie a v sieti rôznych typov buniek sa podieľajú rôzne časti imunitného systému, a to buď priamo kontaktom medzi bunkami alebo uvoľňovaním rôznych cytokínov a produkciou špecifických protilátok (IgE, IgG1, IgG4, IgA ). Ďalšie podrobnosti o mechanizmoch VIT sú znázornené na obrázku 1.
Reakcie špecifických alergénových protilátok
Spočiatku subkutánna VIT je spojená s prechodným skorým zvýšením hladín sérových alergén špecifických IgE (sIgE) a potom nastáva pokles sIgE v priebehu niekoľkých rokov (48–50). AIT je tiež spojená so zvýšením alergén-špecifických protilátok IgA, IgG1 a IgG4, ktoré sa nazývajú blokujúce protilátky (48, 51). Štúdie s aeroalergénmi ukázali významné zvýšenie sérových koncentrácií blokujúcich protilátok až 100-krát v závislosti od času a dávky (52–54). Produkcia blokujúcich protilátok, najmä typu IgG4, môže súťažiť s sIgE o alergén a zabrániť interakcii alergén-sIgE. Blokovanie interakcií alergén-sIgE zabraňuje zosieťovaniu vysokoafinitných IgE receptorov (FcεRI) na bazofiloch a mastocytoch, čo inhibuje degranuláciu týchto buniek a môže zabrániť rozvoju anafylaxie (55, 56). Blokujúce protilátky, IgG/IgG4, inhibujú IgE-uľahčenú prezentáciu alergénu T bunkám blokovaním nízkoafinitných receptorov (FcgRIIb) na B bunkách a zastavujú alergénom indukované zvýšenie produkcie pamäťových IgE (54, 57, 58). Jeden z hlavných cytokínov produkovaných T regs, IL-10, sa nielen podieľa na supresii alergén-špecifických T efektorových buniek počas AIT, ale tiež inhibuje produkciu celkového IgE a špecifického IgE, pričom zvyšuje hladiny IgG4 (59 , 60). U nealergických včelárov sú sérové koncentrácie IgG4 takmer tisíckrát vyššie ako sérové špecifické hladiny IgE. Koncentrácie IgG4 v sére úzko korelujú s počtom bodnutí za rok a časom stráveným vo včelárstve (31, 61). Na zvieracom modeli alergie na včelí jed bol hlavný alergén na včelí jed PLA2 injikovaný do inguinálnych lymfatických uzlín a bola pozorovaná alergén-špecifická IgG odpoveď s produkciou IgG2a (62). V ďalšej štúdii peptidovej VIT sa zistilo zníženie alergén-špecifického IgE a zvýšenie špecifického IgG2a, pričom obe mali preventívnu funkciu proti alergénom indukovanej anafylaxii (63).
Po ukončení AIT pre alergiu na trávový peľ sa ukázalo, že blokujúce hladiny IgG/IgG4 sa v priebehu 1 roka znížili takmer o 80–90 %. Na rozdiel od toho, sérová inhibičná aktivita IgE spojená s IgG pretrvávala niekoľko rokov a korelovala s klinickou účinnosťou (64). To naznačuje, že presnejším meradlom klinickej účinnosti môže byť skôr funkčná aktivita blokujúcich protilátok než ich hladiny a zdá sa, že úzko koreluje s dlhodobou imunitnou toleranciou (54). To však nemusí platiť pre imunoterapiu včelím jedom, kde hoci bola úspešná desenzibilizácia sprevádzaná zvýšením inhibičnej aktivity IgG4 aj IgE, zvýšené špecifické hladiny IgG4 aj inhibičná aktivita IgE-FAB sa vrátili na základnú hodnotu v priebehu mesiacov po prerušení liečby VIT a ďalšie sledovanie odhalili trvalejší pokles hladín IgE špecifických pre jed (65), čo predstavuje predpokladaný alternatívny mechanizmus predĺženej ochrany po vysadení IgG. Podporuje to aj pozorovanie nízkych/neprítomných hladín IgE u tolerantných včelárov (66).
Odpovede efektorových buniek
VIT prejavuje svoje pôsobenie prostredníctvom komplexných imunologických mechanizmov. Počiatočný mechanizmus účinku pozorovaný na efektorových bunkách je desenzibilizácia žírnych buniek a bazofilov (9). Počet týchto buniek sa počas imunoterapie jedom znižuje a navyše ich prahy uvoľňovania cytokínov sa časom zvyšujú. Počas rýchlej VIT na začiatku liečby dochádza k poklesu počtu bazofilov v periférnej krvi a tiež k zníženiu produkcie a uvoľňovania cytokínov odvodených od bazofilov, ako sú IL-4 a IL-13 (67). Bazálna hladina tryptázy v sére, ktorá je markerom záťaže žírnych buniek a funkcie žírnych buniek, sa v priebehu VIT časom znižuje (68). U pacientov s VIT sa už predtým preukázala supresia povrchových antigénov na krvných bazofiloch (69). Okrem zmien pozorovaných v bazofilných povrchových antigénoch sa u pacientov s VIT znížilo aj množstvo histamínu uvoľneného z bazofilov po stimulácii bodnutím v závislosti od ich klinickej reaktivity (70). Reaktivita bazofilov hodnotená prietokovým cytometrickým hodnotením expresie CD63 sa ukázala ako spoľahlivý diagnostický test na diagnostiku alergie na jed blanokrídleho hmyzu (71). Senzitivita bazofilov, dávka, pri ktorej nastane polovica maximálnej bazofilnej odpovede, bola navrhnutá na monitorovanie VIT (72–74). Nový spôsob funkčného testu, ktorý meria intracelulárne farbenie daimínoxidázy konjugovanej s fykoerytrínom (DAO), bol overený na detekciu množstva histamínu uvoľneného z bazofilov. Po stimulácii alergénom sa intracelulárne hladiny DAO znižujú úmerne s uvoľneným intracelulárnym histamínom. Toto zníženie sa preukázalo u pacientov liečených vespula VIT, čo je dôležité pre zvýšenie prahu pre jed na vyvolanie anafylaktickej reakcie u pacientov s VIT (75). U pacientov po VIT klesá nielen vopred vytvorené uvoľňovanie mediátorov, ale aj produkcia a uvoľňovanie novovytvorených mediátorov, ako je leukotrién C4 v krvných bazofiloch (76).
Po rýchlej VIT bol v kultúrach mononukleárnych buniek periférnej krvi (PBMC) hlásený pokles produkcie proteínu exprimovaného a secernovaného T-bunkami (RANTES), IL-8 a monocytového chemoatraktantového proteínu 1 (MCP-1) na úrovni proteínu a mRNA. úrovne (77).
Počas počiatočných fáz VIT nie sú úplne pochopené mechanizmy, ktoré spúšťajú desenzibilizáciu. V roku 2010 Bussmann a spol. vykonali štúdiu na pacientoch s náhlou VIT. Analyzovali hladiny expresie rôznych tolerogénnych markerov na hladinách proteínov a mRNA počas prvých 5 dní VIT. Pozorovali výraznú degradáciu tryptofánu, ktorá je spojená so supresiou reakcií T buniek a indukciou tolerancie; zvýšené ILT3 a ILT4, ktoré sú inhibičnými receptormi pre monocyty, a produkcia IL-10 CD3+ T buniek a monocytov nasledovaná zvýšenými hladinami IL-10 v sére, čo je dôležitý regulačný interleukín na potlačenie alergénom indukovaných reakcií (78).
V štúdiách s aeroalergénmi sa ukázalo, že AIT inhibuje skorú a neskorú fázu alergických reakcií v miestach alergických tkanív prostredníctvom supresie niekoľkých cytokínov a zníženia počtu eozinofilov, mastocytov a bazofilov (54). Tieto informácie naznačujú, že alergénová imunoterapia je účinná na systémovej aj lokálnej úrovni. Podobné mechanizmy sa pravdepodobne uplatňujú aj pri VIT.
Reakcie T a B buniek počas VIT
Ukázalo sa, že rozvoj imunitnej tolerancie počas VIT súvisí s modifikáciou odpovedí T a B buniek (79). Počas VIT dochádza k posunu Th2 na Th1 a pozoruje sa zvýšenie hladín interferónu gama (IFN-γ) paralelne s poklesom IL-4 a IL-13 v plnej krvi (80, 81). Th2 odpovede počas VIT sú znížené a dochádza tiež k zvýšeniu počtu a funkcií Treg buniek (80, 82). Treg bunky sú rozdelené do 2 podskupín ako prirodzené regulačné T (nTreg) bunky, ktoré sú charakterizované transkripčným faktorom forkhead box P3 (FOXP3), a indukovateľné regulačné T (iTreg) bunky, ako sú IL-10 produkujúce Tr1 bunky a TGF- b produkujúce bunky TH3 (83–85). IL-10 hrá inhibičnú úlohu v B bunkách blokovaním B7/CD28 dráhy. To má za následok supresívny účinok na dozrievanie dendritických buniek a na expresiu MHC triedy II a kostimulačných ligandov (86). TGF-b downreguluje expresiu FceRI na Langerhansových bunkách a tiež upreguluje FOXP3 a RUNX a napomáha expresii CTLA-4 na T bunkách (87, 88).
U včelárov Treg bunky produkujúce IL-10 inhibujú proliferáciu PLA-špecifických efektorových T buniek krátko po začiatku sezóny včelieho jedu. Tento supresívny účinok možno zvrátiť blokovaním receptorov CTLA-4, PD-1 a IL-10 (89). Okrem toho indukcia enzýmu indolamín 2,3-dioxygenázy v dendritických bunkách účinkom Tregs spôsobuje transformáciu zápalových dendritických buniek na regulačné dendritické bunky (90). Podobným spôsobom, počas VIT, je proliferácia Treg buniek typu Tr1 prominentná a antigén-špecifické proliferatívne a cytokínové odpovede proti hlavnému alergénu včelieho jedu, fosfolipáze A2 (PLA), boli do konca prvého týždňa VIT významne potlačené. (91). Alergénom indukovaná sekrécia Th2 cytokínov, ako je IL-4, IL-5 a IL-13, bola zrušená (92). Okrem produkcie IL-10 môžu Treg bunky tiež potláčať imunitné reakcie prostredníctvom interakcií medzi bunkami.
Úloha zvýšených hladín IL-10 je významná pri rozvoji klinickej a imunologickej tolerancie počas VIT. Blokovanie IL-10 v PBMC rekonštituuje špecifické proliferatívne a cytokínové reakcie. Túto situáciu možno pozorovať aj u včelárov, ktorí dostali viacnásobné včelie bodnutie (91). Prítomnosť zvýšeného počtu CD4+CD25+FOXP3+ Treg buniek v cieľovom orgáne, nosovej sliznici, po alergénovej imunoterapii trávovým peľom naznačuje, že Treg bunky hrajú dôležitú úlohu pri rozvoji alergén-špecifickej imunitnej tolerancie (93). Podobným spôsobom sa zistilo, že VIT súvisí s progresívnou expanziou počtu cirkulujúcich CD4+CD25+FOXP3+ Treg buniek (94). Počas všetkých typov AIT bola hlásená odchýlka smerom k regulačnej/supresorovej T bunkovej odpovedi (95). V štúdii Nassera et al sa alergénom indukované zmeny v cytokínovej mRNA a bunkových profiloch z kožných biopsií porovnávali pred a 3 mesiace po osej VIT. Došlo k významnému poklesu IL-4 mRNA a zvýšeniu IL-10+ buniek. Okrem toho bol tiež pozorovaný trend k zvýšeniu IL-10 mRNA (96). V ďalšej štúdii Schuerwegha et al sa hodnotil účinok VIT na CD4+CD8+ T lymfocyty pred VIT, na konci 5-dňovej semi-rush VIT a 6 mesiacov počas VIT. Významný pokles počtu CD4+ a CD8+ T buniek produkujúcich IL v porovnaní s bunkami produkujúcimi cytokíny pred VIT bol pozorovaný na konci 5-dňovej semi-rush VIT. Po 6 mesiacoch VIT sa zistilo vyššie množstvo CD4+CD8+ T lymfocytov produkujúcich IL-2 a IFN-γ, čo potvrdzuje posun z imunitnej odchýlky typu Th2 na Th1 (81). Hladiny IL-10 v sére sa začali zvyšovať od druhého dňa VIT (78) a na 28. deň liečby sa v alergén-špecifických T bunkách objavil desenzibilizovaný stav spojený s priamymi supresívnymi účinkami IL-10 (94).
T folikulárne pomocné bunky (TFH) sú definované povrchovým receptorom CXCR5+ a pomáhajú pri dozrievaní B-buniek a pri prepínaní triedy imunoglobulínov. CXCR5+ FoxP3+ Treg bunky sú podskupinou Tregs, nazývaných ako folikulárne regulačné T (TFR) bunky, ktoré sú schopné potlačiť reakcie T- a B-buniek migráciou do germinálnych centier lymfatických uzlín (97, 98). Štúdia imunoterapie trávovým peľom preukázala významný pokles počtu pamäťových TFH buniek po imunoterapii (99). Okrem toho sa zistilo, že bunky TFR produkujú viac IL-10 v porovnaní s bunkami TFH. Plasticita medzi TFH a TFR bunkami bola preukázaná v tej istej štúdii, čo naznačuje, že TFR bunky môžu hrať dôležitú úlohu pri potláčaní TH2 odpovedí a produkcie alergénovo špecifických IgE počas imunoterapie (99). Je pravdepodobné, že počas imunoterapie jedom sú prítomné podobné mechanizmy TFR a TFH buniek ako pri imunoterapii peľom tráv.
Nedávno boli u včelárov tolerujúcich včelí jed a u pacientov, ktorým sa podávala VIT, identifikované bunky Breg špecifické pre alergén vylučujúce IL-10 (100). Breg bunky sú charakterizované ako CD73− CD25+CD71+ B bunky, ktoré sú schopné potláčať CD4+ T bunky špecifické pre včelí jed a schopné produkovať alergén špecifické IgG4 protilátky po včelej VIT (100). Okrem toho môžu Breg bunky tiež preukázať svoju inhibičnú kapacitu produkciou IL-35 a TGF-beta (101). Okrem Treg a B reg buniek sa ukázalo, že prirodzené zabíjačské regulačné bunky vylučujúce IL-10 potláčajú alergénom stimulovanú proliferáciu T buniek u ľudí a môžu byť dôležité pri indukcii tolerancie ako iné typy regulačných buniek (102).
Vrodené lymfoidné bunky a alergénová imunoterapia
Účinok alergénovej imunoterapie na vrodené lymfoidné bunky, ILC typu 2, bol študovaný pri alergii na peľ tráv v periférnej krvi. AIT potlačila sezónne zvýšenia ILC2 u pacientov liečených imunoterapiou v porovnaní s neliečenými kontrolami (103). Pokles ILC2 koreloval so symptómami, ktoré sami hlásili. Okrem toho sa tiež znížil podiel IL-13+ ILC2. V ďalšej štúdii sezónnych astmatických pacientov Lombardi et al nedokázali preukázať žiadnu zmenu v počte ILC2 počas imunoterapie, čo bolo vysvetlené nesezónnymi meraniami, zatiaľ čo pacienti boli asymptomatickí (104). Doteraz neexistuje dôkaz, že imunoterapia má nejaký účinok na cytokíny odvodené z epitelu, ako sú IL-25, IL-33 a TSLP, ktoré majú regulačné účinky na lokálny zápal typu 2 a ILC (54).
Histamín a histamínové receptory na VIT
Počas VIT sa v priebehu niekoľkých dní alebo dokonca hodín rozvinie skorá desenzibilizácia v závislosti od typu použitého imunoterapeutického protokolu, ako je náhly a ultrarýchly typ VIT. Dochádza k poklesu počtu bazofilov, vopred vytvorených mediátorov a uvoľňovania mediátorov v závislosti od času (75, 105, 106). Medzi štyrmi rôznymi typmi histamínových receptorov hrá dôležitú úlohu histamínový receptor typu 2 (HR2) s toleranciou periférneho antigénu (89). Potlačenie bazofilov začína aktiváciou histamínových receptorov typu 2 (HR2). H2R znižuje alergénom indukovanú degranuláciu bazofilov sprostredkovanú FceRI a uvoľňovanie mediátora (107). HR2 sa podieľa hlavne na tolerogénnych imunitných odpovediach. Je upregulovaný v Th2 bunkách a oba potláčajú alergénom stimulované reakcie T buniek a zvyšujú produkciu IL-10 u včelárov (89), čo vyvoláva rozvoj periférnej tolerancie (76, 108, 109). Histamín prostredníctvom HR2 indukuje produkciu IL-10 dendritickými bunkami a Th2 bunkami (110); zvyšuje supresívny účinok TGF-b na T bunky (111) a znižuje produkciu IL-4 a IL-13, čo sú hlavné cytokíny typu Th2 (112).
Účinnosť a bezpečnosť imunoterapie jedom
Účinnosť VIT sa pohybuje od 77 do 84 % pre včelu medonosnú a od 91 do 96 % pre jed osy (9–11). Niektoré faktory, ako napríklad vyššie množstvo alergénu preneseného počas každej reakcie na bodnutie, konzistencia včelieho bodnutia, rôznorodosť vzoru senzibilizácie včelieho jedu, patria medzi navrhované faktory, ktoré môžu vysvetliť nižšiu mieru úspešnosti spojenú s VIT včelou (9). Zistilo sa, že VIT je účinná dokonca aj po fáze nahromadenia a v jednej štúdii s jedom včiel bola hlásená 89 % úspešnosť pri stimulácii bodnutím 1 týždeň po dosiahnutí udržiavacej dávky (113).
Vzhľadom na imunoterapiu včelím jedom môže použitie vzorcov senzibilizácie s rozlíšenými komponentmi pomôcť zvýšiť úspešnosť liečby. Niektorí pacienti sú senzibilizovaní predovšetkým na Api m 10, čo je nedostatočne zastúpený alergén v niektorých prípravkoch VIT, ktorý môže spôsobiť zlyhanie liečby (114, 115).
Počas fázy budovania VIT, ak je SSR problémom dosiahnuť udržiavaciu dávku, môže byť alternatívou premedikácia omalizumabom, anti-IgE protilátkou (116).
Na prevenciu zlyhania liečby je dôležitá aj dávka jedu použitá počas VIT. Zvyčajne na ochranu postačuje udržiavacia dávka 100 μg jedu počas VIT (9). Keď sú rizikové faktory vysoké, ako u včelárov, vyššia dávka jedu dáva lepšie výsledky (117). Ak sa systémová reakcia rozvinie po terénnom bodnutí alebo stimulácii bodnutím počas konvenčnej dávky 100 μg, odporúča sa vyššia dávka, 200 μg (118).
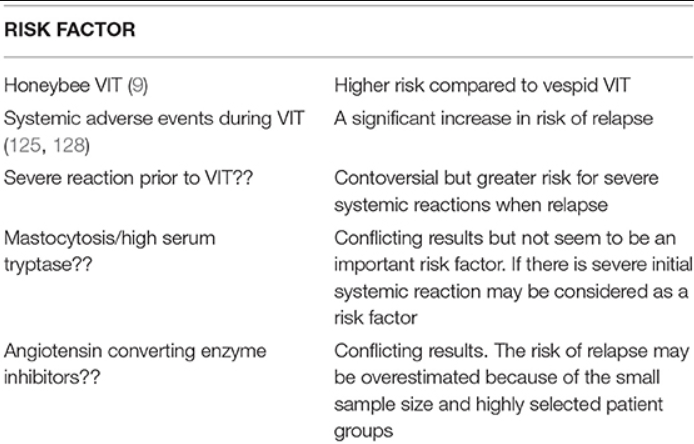
Miera relapsov až do 10 % bola hlásená za 1–5 rokov po ukončení vespula VIT a častejšie pri VIT včiel (9). V jednej štúdii bola 3–5 rokov po ukončení VIT hlásená miera relapsov 7,5 % pre vespula VIT a 15,8 % pre VIT včiel (17). U detí má VIT lepšiu prognózu v porovnaní s dospelými a len u 5 % detí sa vyvinú stredne ťažké až ťažké systémové reakcie na bodnutie do 20 rokov sledovania po ukončení VIT (15).
Trvanie VIT je dôležité pre účinnosť. Jeden rok liečby zlyhal u takmer jednej štvrtiny pacientov, keď ich štípalo 3–4 roky po VIT (119). Zdá sa, že štúdie s najmenej 5-ročnou liečbou preukazujú lepšiu ochranu v porovnaní s 3-ročnou liečbou (120). Lerch et al preukázali nižšie miery systémových reakcií u pacientov, ktorí dostávali VIT ≥ 50 mesiacov v porovnaní s pacientmi liečenými 33–49 mesiacov (5 vs. 18 %) (17). V inej štúdii Golden et al. hodnotili pacientov liečených VIT aspoň 5 rokov a zistili mieru systémovej reakcie 9,5 % do 5 rokov po ukončení VIT (121).
Nedávna metaanalýza zahŕňajúca päť systematických prehľadov, päť RCT, tri kontrolované štúdie (pred a po) a štyri série prípadov ukázali, že VIT významne znížila riziko závažných systémových reakcií (OR = 0,08, 95 % CI 0,03–0,26); zlepšená kvalita života (rozdiel rizika: 1,41, 95 % CI 1,04–1,79) a môže byť nákladovo efektívna u tých, ktorí zaznamenali opakované systémové reakcie a zhoršenú kvalitu života (122).
Nežiaduce udalosti sú počas VIT zvyčajne mierne. U pacientov so systémovými reakciami dochádza k relapsu oveľa častejšie v porovnaní s pacientmi, ktorí ho nemali (16,4–38 % vs. 5,4–8 %) (11).
Niektoré z predchádzajúcich rizikových faktorov systémových nežiaducich udalostí počas VIT sa už nepovažujú za dôležité rizikové faktory. K týmto starším rizikovým faktorom patrí mastocytóza, ACE inhibítory, užívanie betablokátorov, vysoké hladiny špecifických IgE a pozitivita kožných prick testov pri nízkej záťaži testov (9). Vysoké bazálne hladiny tryptázy pri alergii na vespid môžu byť rizikovým faktorom pre systémové nežiaduce udalosti pri VIT (12), ale nie pri alergii na včelí jed (123).
Rizikové faktory relapsu SSR po ukončení VIT sú uvedené v tabuľke 2.
V súčasnosti je zlatým štandardom na identifikáciu účinnosti VIT a na odlíšenie respondentov od pacientov, ktorí nereagujú, test bodnutia. Metódy používané na monitorovanie VIT sú uvedené v tabuľke 3.

Záver
Alergia na hmyzí jed je jednou z najčastejších príčin anafylaxie u ľudí a ide o zdravotnú pohotovosť. V súčasnosti neexistuje žiadny biomarker na predpovedanie rizika anafylaxie. VIT je najúčinnejšou liečbou na prevenciu SSR po bodnutí a znižuje riziko anafylaxie. Stále však treba odpovedať na niektoré otázky, ako sú náklady na účinnosť, vplyv na kvalitu života, trvanie liečby, optimálna dávka a spôsoby hodnotenia. Rozvoj periférnej tolerancie je hlavným mechanizmom počas VIT, ktorý je riadený T regulačnými bunkami. T regs produkujú IL-10 a potláčajú Th2 imunitu a imunitné reakcie sa posúvajú smerom k zápalu typu Th1. Blokujúce protilátky IgG1/IgG4 inhibujú reakcie závislé od IgE na žírnych bunkách, bazofiloch a B bunkách. Na rozvoji dlhodobej imunitnej tolerancie sa podieľa aj niekoľko ďalších mechanizmov, ako sú epitelové bunky, niekoľko cytokínov, dendritické bunky, ILC2 a B regulačné bunky.
Faktory, akými sú zvyšovanie vedomostí o rizikových faktoroch anafylaxie jedom, lepšia edukácia pacientov, vývoj efektívnejších produktov VIT s menšími vedľajšími účinkami a nájdenie účinných biomarkerov na predpovedanie budúcich systémových reakcií na individuálnej úrovni, výrazne zlepšia starostlivosť o pacienta.
Autorské príspevky
USA vykonali rešerš literatúry, pripravili obrázok a tabuľky a napísali článok pod dohľadom SD. SD naplánoval celý text, prečítal rukopis, urobil potrebné opravy a podrobne diskutoval o mechanizmoch imunoterapie.
Vyhlásenie o konflikte záujmov
SD vykazuje granty od ALK, Dánsko, osobné poplatky od Anergis, Švajčiarsko, osobné poplatky od Biomay, Rakúsko, osobné poplatky od Allergy Therapeutics, UK, osobné poplatky od ALK, Horsholm, Dánsko, osobné poplatky od Allergy Therapeutics, mimo zaslanej práce.
Zostávajúci autor vyhlasuje, že výskum bol vykonaný bez akýchkoľvek obchodných alebo finančných vzťahov, ktoré by mohli byť chápané ako potenciálny konflikt záujmov.
Referencie
1. Krishna MT, Ewan PW, Diwakar L, Durham SR, Frew AJ, Leech SC, et al. Diagnosis and management of hymenoptera venom allergy: British society for allergy and clinical immunology (BSACI) guidelines. Clin Exp Allergy. (2011) 41:1201–20. doi: 10.1111/j.1365-2222.2011.03788.x
PubMed Abstract | CrossRef Full Text | Google Scholar
2. Tan JW, Campbell DE. Insect allergy in children. J Paediatr Child Health. (2013) 49:E381–7. doi: 10.1111/jpc.12178
PubMed Abstract | CrossRef Full Text | Google Scholar
3. Hoffman DR, Jacobson RS. Allergens in hymenoptera venom XII: how much protein is in a sting? Ann Allergy. (1984) 52:276–8.
PubMed Abstract | Google Scholar
4. Ellis AK, Day JH. Clinical reactivity to insect stings. Curr Opin Allergy Clin Immunol. (2005) 5:349–54. doi: 10.1097/01.all.0000174159.55756.58
PubMed Abstract | CrossRef Full Text | Google Scholar
5. Mauriello PM, Barde SH, Georgitis JW, Reisman RE. Natural history of large local reactions from stinging insects. J Allergy Clin Immunol. (1984) 74(4 Pt 1):494–8. doi: 10.1016/0091-6749(84)90384-1
PubMed Abstract | CrossRef Full Text | Google Scholar
6. Ludman SW, Boyle RJ. Stinging insect allergy: current perspectives on venom immunotherapy. J Asthma Allergy. (2015) 8:75–86. doi: 10.2147/JAA.S62288
PubMed Abstract | CrossRef Full Text | Google Scholar
7. Bilo BM, Bonifazi F. Epidemiology of insect-venom anaphylaxis. Curr Opin Allergy Clin Immunol. (2008) 8:330–7. doi: 10.1097/ACI.0b013e32830638c5
PubMed Abstract | CrossRef Full Text | Google Scholar
8. Jennings A, Duggan E, Perry IJ, Hourihane JO. Epidemiology of allergic reactions to hymenoptera stings in Irish school children. Pediatr Allergy Immunol. (2010) 21:1166–70. doi: 10.1111/j.1399-3038.2010.01054.x
PubMed Abstract | CrossRef Full Text | Google Scholar
9. Sturm GJ, Varga EM, Roberts G, Mosbech H, Bilo MB, Akdis CA, et al. EAACI guidelines on allergen immunotherapy: Hymenoptera venom allergy. Allergy. (2018) 73:744–64. doi: 10.1111/all.13262
PubMed Abstract | CrossRef Full Text | Google Scholar
10. Muller U, Helbling A, Berchtold E. Immunotherapy with honeybee venom and yellow jacket venom is different regarding efficacy and safety. J Allergy Clin Immunol. (1992) 89:529–35. doi: 10.1016/0091-6749(92)90319-W
PubMed Abstract | CrossRef Full Text | Google Scholar
11. Rueff F, Vos B, Oude Elberink J, Bender A, Chatelain R, Dugas-Breit S, et al. Predictors of clinical effectiveness of Hymenoptera venom immunotherapy. Clin Exp Allergy. (2014) 44:736–46. doi: 10.1111/cea.12275
PubMed Abstract | CrossRef Full Text | Google Scholar
12. Rueff F, Przybilla B, Bilo MB, Muller U, Scheipl F, Aberer W, et al. Predictors of side effects during the buildup phase of venom immunotherapy for Hymenoptera venom allergy: the importance of baseline serum tryptase. J Allergy Clin Immunol. (2010) 126:105–11.e5. doi: 10.1016/j.jaci.2010.04.025
PubMed Abstract | CrossRef Full Text | Google Scholar
13. Mosbech H, Muller U. Side-effects of insect venom immunotherapy: results from an EAACI multicenter study. European Academy of Allergology and Clinical Immunology. Allergy. (2000) 55:1005–10. doi: 10.1034/j.1398-9995.2000.00587.x
PubMed Abstract | CrossRef Full Text | Google Scholar
14. Stoevesandt J, Hain J, Stolze I, Kerstan A, Trautmann A. Angiotensin-converting enzyme inhibitors do not impair the safety of Hymenoptera venom immunotherapy build-up phase. Clin Exp Allergy. (2014) 44:747–55. doi: 10.1111/cea.12276
CrossRef Full Text | Google Scholar
15. Golden DB, Kagey-Sobotka A, Norman PS, Hamilton RG, Lichtenstein LM. Outcomes of allergy to insect stings in children, with and without venom immunotherapy. N Engl J Med. (2004) 351:668–74. doi: 10.1056/NEJMoa022952
PubMed Abstract | CrossRef Full Text | Google Scholar
16. Reisman RE. Duration of venom immunotherapy: relationship to the severity of symptoms of initial insect sting anaphylaxis. J Allergy Clin Immunol. (1993) 92:831–6. doi: 10.1016/0091-6749(93)90060-S
PubMed Abstract | CrossRef Full Text | Google Scholar
17. Lerch E, Muller UR. Long-term protection after stopping venom immunotherapy: results of re-stings in 200 patients. J Allergy Clin Immunol. (1998) 101:606–12. doi: 10.1016/S0091-6749(98)70167-8
PubMed Abstract | CrossRef Full Text | Google Scholar
18. Novembre E, Cianferoni A, Bernardini R, Veltroni M, Ingargiola A, Lombardi E, et al. Epidemiology of insect venom sensitivity in children and its correlation to clinical and atopic features. Clin Exp Allergy. (1998) 28:834–8. doi: 10.1046/j.1365-2222.1998.00313.x
PubMed Abstract | CrossRef Full Text | Google Scholar
19. Grigoreas C, Galatas ID, Kiamouris C, Papaioannou D. Insect-venom allergy in Greek adults. Allergy. (1997) 52:51–7. doi: 10.1111/j.1398-9995.1997.tb02545.x
PubMed Abstract | CrossRef Full Text | Google Scholar
20. Incorvaia C, Mauro M, Pastorello EA. Hymenoptera stings in conscripts. Allergy. (1997) 52:680–1. doi: 10.1111/j.1398-9995.1997.tb01056.x
PubMed Abstract | CrossRef Full Text | Google Scholar
21. Fernandez J, Blanca M, Soriano V, Sanchez J, Juarez C. Epidemiological study of the prevalence of allergic reactions to Hymenoptera in a rural population in the Mediterranean area. Clin Exp Allergy. (1999) 29:1069–74. doi: 10.1046/j.1365-2222.1999.00614.x
PubMed Abstract | CrossRef Full Text | Google Scholar
22. Navarro LA, Pelaez A, de la Torre F, Tenias Burillo JM, Megias J, Martinez I. Epidemiological factors on hymenoptera venom allergy in a Spanish adult population. J Investig Allergol Clin Immunol. (2004) 14:134–41.
PubMed Abstract | Google Scholar
23. Fernandez J, Soriano V, Mayorga L, Mayor M. Natural history of Hymenoptera venom allergy in Eastern Spain. Clin Exp Allergy. (2005) 35:179–85. doi: 10.1111/j.1365-2222.2005.02169.x
PubMed Abstract | CrossRef Full Text | Google Scholar
24. Bilo MB, Cichocka-Jarosz E, Pumphrey R, Oude-Elberink JN, Lange J, Jakob T, et al. Self-medication of anaphylactic reactions due to Hymenoptera stings-an EAACI task force consensus statement. Allergy. (2016) 71:931–43. doi: 10.1111/all.12908
PubMed Abstract | CrossRef Full Text | Google Scholar
25. Muller UR. Bee venom allergy in beekeepers and their family members. Curr Opin Allergy Clin Immunol. (2005) 5:343–7. doi: 10.1097/01.all.0000173783.42906.95
PubMed Abstract | CrossRef Full Text | Google Scholar
26. Richter AG, Nightingale P, Huissoon AP, Krishna MT. Risk factors for systemic reactions to bee venom in British beekeepers. Ann Allergy Asthma Immunol. (2011) 106:159–63. doi: 10.1016/j.anai.2010.11.005
PubMed Abstract | CrossRef Full Text | Google Scholar
27. Worm M, Moneret-Vautrin A, Scherer K, Lang R, Fernandez-Rivas M, Cardona V, et al. First European data from the network of severe allergic reactions (NORA). Allergy. (2014) 69:1397–404. doi: 10.1111/all.12475
PubMed Abstract | CrossRef Full Text | Google Scholar
28. Prado M, Quiros D, Lomonte B. Mortality due to Hymenoptera stings in Costa Rica, 1985-2006. Rev Panam Salud Publica. (2009) 25:389–93. doi: 10.1590/S1020-49892009000500002
PubMed Abstract | CrossRef Full Text | Google Scholar
29. Barnard JH. Studies of 400 Hymenoptera sting deaths in the United States. J Allergy Clin Immunol. (1973) 52:259–64. doi: 10.1016/0091-6749(73)90044-4
PubMed Abstract | CrossRef Full Text | Google Scholar
30. Turner PJ, Gowland MH, Sharma V, Ierodiakonou D, Harper N, Garcez T, et al. Increase in anaphylaxis-related hospitalizations but no increase in fatalities: an analysis of United Kingdom national anaphylaxis data, 1992-2012. J Allergy Clin Immunol. (2015) 135:956–63.e1. doi: 10.1016/j.jaci.2014.10.021
PubMed Abstract | CrossRef Full Text | Google Scholar
31. Bilo BM, Rueff F, Mosbech H, Bonifazi F, Oude-Elberink JN, EAACI Interest Group on Insect Venom Hypersensitivity. Diagnosis of Hymenoptera venom allergy. Allergy. (2005) 60:1339–49. doi: 10.1111/j.1398-9995.2005.00963.x
PubMed Abstract | CrossRef Full Text | Google Scholar
32. Clark S, Camargo CA Jr. Epidemiology of anaphylaxis. Immunol Allergy Clin North Am. (2007) 27:145–63. doi: 10.1016/j.iac.2007.03.002
PubMed Abstract | CrossRef Full Text | Google Scholar
33. Rueff F, Przybilla B, Bilo MB, Muller U, Scheipl F, Aberer W, et al. Predictors of severe systemic anaphylactic reactions in patients with Hymenoptera venom allergy: importance of baseline serum tryptase-a study of the European Academy of Allergology and Clinical Immunology Interest Group on Insect Venom Hypersensitivity. J Allergy Clin Immunol. (2009) 124:1047–54. doi: 10.1016/j.jaci.2009.08.027
PubMed Abstract | CrossRef Full Text | Google Scholar
34. Arzt L, Bokanovic D, Schwarz I, Schrautzer C, Massone C, Horn M, et al. Hymenoptera stings in the head region induce impressive, but not severe sting reactions. Allergy. (2016) 71:1632–4. doi: 10.1111/all.12967
CrossRef Full Text | Google Scholar
35. Brockow K, Jofer C, Behrendt H, Ring J. Anaphylaxis in patients with mastocytosis: a study on history, clinical features and risk factors in 120 patients. Allergy. (2008) 63:226–32. doi: 10.1111/j.1398-9995.2007.01569.x
PubMed Abstract | CrossRef Full Text | Google Scholar
36. Castells MC, Hornick JL, Akin C. Anaphylaxis after hymenoptera sting: is it venom allergy, a clonal disorder, or both? J Allergy Clin Immunol Pract. (2015) 3:350–5. doi: 10.1016/j.jaip.2015.03.015
PubMed Abstract | CrossRef Full Text | Google Scholar
37. Bonadonna P, Bonifacio M, Lombardo C, Zanotti R. Hymenoptera allergy and mast cell activation syndromes. Curr Allergy Asthma Rep. (2016) 16:5. doi: 10.1007/s11882-015-0582-5
PubMed Abstract | CrossRef Full Text | Google Scholar
38. Stoevesandt J, Hain J, Kerstan A, Trautmann A. Over- and underestimated parameters in severe Hymenoptera venom-induced anaphylaxis: cardiovascular medication and absence of urticaria/angioedema. J Allergy Clin Immunol. (2012) 130:698–704.e1. doi: 10.1016/j.jaci.2012.03.024
PubMed Abstract | CrossRef Full Text | Google Scholar
39. Mueller UR. Cardiovascular disease and anaphylaxis. Curr Opin Allergy Clin Immunol. (2007) 7:337–41. doi: 10.1097/ACI.0b013e328259c328
PubMed Abstract | CrossRef Full Text | Google Scholar
40. Valent P, Akin C, Escribano L, Fodinger M, Hartmann K, Brockow K, et al. Standards and standardization in mastocytosis: consensus statements on diagnostics, treatment recommendations and response criteria. Eur J Clin Invest. (2007) 37:435–53. doi: 10.1111/j.1365-2362.2007.01807.x
PubMed Abstract | CrossRef Full Text | Google Scholar
41. Castells M, Metcalfe DD, Escribano L. Diagnosis and treatment of cutaneous mastocytosis in children: practical recommendations. Am J Clin Dermatol. (2011) 12:259–70. doi: 10.2165/11588890-000000000-00000
PubMed Abstract | CrossRef Full Text | Google Scholar
42. Akin C, Valent P. Diagnostic criteria and classification of mastocytosis in 2014. Immunol Allergy Clin North Am. (2014) 34:207–18. doi: 10.1016/j.iac.2014.02.003
PubMed Abstract | CrossRef Full Text | Google Scholar
43. Bonadonna P, Perbellini O, Passalacqua G, Caruso B, Colarossi S, Dal Fior D, et al. Clonal mast cell disorders in patients with systemic reactions to Hymenoptera stings and increased serum tryptase levels. J Allergy Clin Immunol. (2009) 123:680–6. doi: 10.1016/j.jaci.2008.11.018
PubMed Abstract | CrossRef Full Text | Google Scholar
44. Alfaya Arias T, Soriano Gomis V, Soto Mera T, Vega Castro A, Vega Gutierrez JM, Alonso Llamazares A, et al. Hymenoptera allergy committee of the: key issues in hymenoptera venom allergy: an update. J Investig Allergol Clin Immunol. (2017) 27:19–31. doi: 10.18176/jiaci.0123
CrossRef Full Text | Google Scholar
45. Gonzalez de Olano D, de la Hoz Caballer B, Nunez Lopez R, Sanchez Munoz L, Cuevas Agustin M, Dieguez MC, et al. Prevalence of allergy and anaphylactic symptoms in 210 adult and pediatric patients with mastocytosis in Spain: a study of the Spanish network on mastocytosis (REMA). Clin Exp Allergy. (2007) 37:1547–55. doi: 10.1111/j.1365-2222.2007.02804.x
PubMed Abstract | CrossRef Full Text | Google Scholar
46. Alvarez-Twose I, Zanotti R, Gonzalez-de-Olano D, Bonadonna P, Vega A, Matito A, et al. Nonaggressive systemic mastocytosis (SM) without skin lesions associated with insect-induced anaphylaxis shows unique features versus other indolent SM. J Allergy Clin Immunol. (2014) 133:520–8. doi: 10.1016/j.jaci.2013.06.020
PubMed Abstract | CrossRef Full Text | Google Scholar
47. Alvarez-Twose I, Gonzalez de Olano D, Sanchez-Munoz L, Matito A, Esteban-Lopez MI, Vega A, et al. Clinical, biological, and molecular characteristics of clonal mast cell disorders presenting with systemic mast cell activation symptoms. J Allergy Clin Immunol. (2010) 125:1269–78.e2. doi: 10.1016/j.jaci.2010.02.019
PubMed Abstract | CrossRef Full Text | Google Scholar
48. Muller U, Helbling A, Bischof M. Predictive value of venom-specific IgE, IgG and IgG subclass antibodies in patients on immunotherapy with honey bee venom. Allergy. (1989) 44:412–8. doi: 10.1111/j.1398-9995.1989.tb04172.x
PubMed Abstract | CrossRef Full Text | Google Scholar
49. Kemeny DM, Lessof MH, Patel S, Youlten LJ, Williams A, Lambourn E. IgG and IgE antibodies after immunotherapy with bee and wasp venom. Int Arch Allergy Appl Immunol. (1989) 88:247–9. doi: 10.1159/000234799
PubMed Abstract | CrossRef Full Text | Google Scholar
50. Michils A, Baldassarre S, Ledent C, Mairesse M, Gossart B, Duchateau J. Early effect of ultrarush venom immunotherapy on the IgG antibody response. Allergy. (2000) 55:455–62. doi: 10.1034/j.1398-9995.2000.00412.x
PubMed Abstract | CrossRef Full Text | Google Scholar
51. Aalberse RC, Dieges PH, Knul-Bretlova V, Vooren P, Aalbers M, van Leeuwen J. IgG4 as a blocking antibody. Clin Rev Allergy. (1983) 1:289–302.
PubMed Abstract | Google Scholar
52. Wachholz PA, Soni NK, Till SJ, Durham SR. Inhibition of allergen-IgE binding to B cells by IgG antibodies after grass pollen immunotherapy. J Allergy Clin Immunol. (2003) 112:915–22. doi: 10.1016/S0091-6749(03)02022-0
PubMed Abstract | CrossRef Full Text | Google Scholar
53. Oefner CM, Winkler A, Hess C, Lorenz AK, Holecska V, Huxdorf M, et al. Tolerance induction with T cell-dependent protein antigens induces regulatory sialylated IgGs. J Allergy Clin Immunol. (2012) 129:1647–55.e13. doi: 10.1016/j.jaci.2012.02.037
PubMed Abstract | CrossRef Full Text | Google Scholar
54. Shamji MH, Durham SR. Mechanisms of allergen immunotherapy for inhaled allergens and predictive biomarkers. J Allergy Clin Immunol. (2017) 140:1485–98. doi: 10.1016/j.jaci.2017.10.010
PubMed Abstract | CrossRef Full Text | Google Scholar
55. Rispens T, Ooijevaar-de Heer P, Bende O, Aalberse RC. Mechanism of immunoglobulin G4 Fab-arm exchange. J Am Chem Soc. (2011) 133:10302–11. doi: 10.1021/ja203638y
PubMed Abstract | CrossRef Full Text | Google Scholar
56. Flicker S, Valenta R. Renaissance of the blocking antibody concept in type I allergy. Int Arch Allergy Immunol. (2003) 132:13–24. doi: 10.1159/000073260
PubMed Abstract | CrossRef Full Text | Google Scholar
57. van Neerven RJ, Wikborg T, Lund G, Jacobsen B, Brinch-Nielsen A, Arnved J, et al. Blocking antibodies induced by specific allergy vaccination prevent the activation of CD4+ T cells by inhibiting serum-IgE-facilitated allergen presentation. J Immunol. (1999) 163:2944–52.
PubMed Abstract | Google Scholar
58. Wurtzen PA, Lund G, Lund K, Arvidsson M, Rak S, Ipsen H. A double-blind placebo-controlled birch allergy vaccination study II: correlation between inhibition of IgE binding, histamine release and facilitated allergen presentation. Clin Exp Allergy. (2008) 38:1290–301. doi: 10.1111/j.1365-2222.2008.03020.x
PubMed Abstract | CrossRef Full Text | Google Scholar
59. Akdis M, Burgler S, Crameri R, Eiwegger T, Fujita H, Gomez E, et al. Interleukins, from 1 to 37, and interferon-gamma: receptors, functions, and roles in diseases. J Allergy Clin Immunol. (2011) 127:701–21.e1–70. doi: 10.1016/j.jaci.2010.11.050
PubMed Abstract | CrossRef Full Text | Google Scholar
60. Meiler F, Klunker S, Zimmermann M, Akdis CA, Akdis M. Distinct regulation of IgE, IgG4 and IgA by T regulatory cells and toll-like receptors. Allergy. (2008) 63:1455–63. doi: 10.1111/j.1398-9995.2008.01774.x
PubMed Abstract | CrossRef Full Text | Google Scholar
61. Carballido JM, Carballido-Perrig N, Kagi MK, Meloen RH, Wuthrich B, Heusser CH, et al. T cell epitope specificity in human allergic and nonallergic subjects to bee venom phospholipase A2. J Immunol. (1993) 150(8 Pt 1):3582–91.
PubMed Abstract | Google Scholar
62. Martinez-Gomez JM, Johansen P, Erdmann I, Senti G, Crameri R, Kundig TM. Intralymphatic injections as a new administration route for allergen-specific immunotherapy. Int Arch Allergy Immunol. (2009) 150:59–65. doi: 10.1159/000210381
PubMed Abstract | CrossRef Full Text | Google Scholar
63. von Garnier C, Astori M, Kettner A, Dufour N, Heusser C, Corradin G, et al. Allergen-derived long peptide immunotherapy down-regulates specific IgE response and protects from anaphylaxis. Eur J Immunol. (2000) 30:1638–45. doi: 10.1002/1521-4141(200006)30:6<1638::AID-IMMU1638>3.0.CO;2-R
PubMed Abstract | CrossRef Full Text | Google Scholar
64. James LK, Shamji MH, Walker SM, Wilson DR, Wachholz PA, Francis JN, et al. Long-term tolerance after allergen immunotherapy is accompanied by selective persistence of blocking antibodies. J Allergy Clin Immunol. (2011) 127:509–16.e1–5. doi: 10.1016/j.jaci.2010.12.1080
PubMed Abstract | CrossRef Full Text | Google Scholar
65. Varga EM, Francis JN, Zach MS, Klunker S, Aberer W, Durham SR. Time course of serum inhibitory activity for facilitated allergen-IgE binding during bee venom immunotherapy in children. Clin Exp Allergy. (2009) 39:1353–7. doi: 10.1111/j.1365-2222.2009.03303.x
PubMed Abstract | CrossRef Full Text | Google Scholar
66. Varga EM, Kausar F, Aberer W, Zach M, Eber E, Durham SR, et al. Tolerant beekeepers display venom-specific functional IgG4 antibodies in the absence of specific IgE. J Allergy Clin Immunol. (2013) 131:1419–21. doi: 10.1016/j.jaci.2012.08.037
PubMed Abstract | CrossRef Full Text | Google Scholar
67. Plewako H, Wosinska K, Arvidsson M, Bjorkander J, Skov PS, Hakansson L, et al. Basophil interleukin 4 and interleukin 13 production is suppressed during the early phase of rush immunotherapy. Int Arch Allergy Immunol. (2006) 141:346–53. doi: 10.1159/000095461
PubMed Abstract | CrossRef Full Text | Google Scholar
68. Dugas-Breit S, Przybilla B, Dugas M, Arnold A, Pfundstein G, Kuchenhoff H, et al. Serum concentration of baseline mast cell tryptase: evidence for a decline during long-term immunotherapy for Hymenoptera venom allergy. Clin Exp Allergy. (2010) 40:643–9. doi: 10.1111/j.1365-2222.2009.03436.x
PubMed Abstract | CrossRef Full Text | Google Scholar
69. Siegmund R, Vogelsang H, Machnik A, Herrmann D. Surface membrane antigen alteration on blood basophils in patients with Hymenoptera venom allergy under immunotherapy. J Allergy Clin Immunol. (2000) 106:1190–5. doi: 10.1067/mai.2000.110928
PubMed Abstract | CrossRef Full Text | Google Scholar
70. Eberlein-Konig B, Ullmann S, Thomas P, Przybilla B. Tryptase and histamine release due to a sting challenge in bee venom allergic patients treated successfully or unsuccessfully with hyposensitization. Clin Exp Allergy. (1995) 25:704–12. doi: 10.1111/j.1365-2222.1995.tb00007.x
PubMed Abstract | CrossRef Full Text | Google Scholar
71. Sturm GJ, Bohm E, Trummer M, Weiglhofer I, Heinemann A, Aberer W. The CD63 basophil activation test in Hymenoptera venom allergy: a prospective study. Allergy. (2004) 59:1110–7. doi: 10.1111/j.1398-9995.2004.00400.x
PubMed Abstract | CrossRef Full Text | Google Scholar
72. Mikkelsen S, Bibby BM, Dolberg MK, Dahl R, Hoffmann HJ. Basophil sensitivity through CD63 or CD203c is a functional measure for specific immunotherapy. Clin Mol Allergy. (2010) 8:2. doi: 10.1186/1476-7961-8-2
PubMed Abstract | CrossRef Full Text | Google Scholar
73. Zitnik SE, Vesel T, Avcin T, Silar M, Kosnik M, Korosec P. Monitoring honeybee venom immunotherapy in children with the basophil activation test. Pediatr Allergy Immunol. (2012) 23:166–72. doi: 10.1111/j.1399-3038.2011.01233.x
PubMed Abstract | CrossRef Full Text | Google Scholar
74. Erzen R, Kosnik M, Silar M, Korosec P. Basophil response and the induction of a tolerance in venom immunotherapy: a long-term sting challenge study. Allergy. (2012) 67:822–30. doi: 10.1111/j.1398-9995.2012.02817.x
PubMed Abstract | CrossRef Full Text | Google Scholar
75. Nullens S, Sabato V, Faber M, Leysen J, Bridts CH, De Clerck LS, et al. Basophilic histamine content and release during venom immunotherapy: insights by flow cytometry. Cytometry B Clin Cytom. (2013) 84:173–8. doi: 10.1002/cyto.b.21084
PubMed Abstract | CrossRef Full Text | Google Scholar
76. Jutel M, Muller UR, Fricker M, Rihs S, Pichler WJ, Dahinden C. Influence of bee venom immunotherapy on degranulation and leukotriene generation in human blood basophils. Clin Exp Allergy. (1996) 26:1112–8. doi: 10.1046/j.1365-2222.1996.d01-259.x
PubMed Abstract | CrossRef Full Text | Google Scholar
77. Akoum H, Duez C, Vorng H, Fahy O, Wallaert B, Tonnel AB, et al. Early modifications of chemokine production and mRNA expression during rush venom immunotherapy. Cytokine. (1998) 10:706–12. doi: 10.1006/cyto.1998.0348
PubMed Abstract | CrossRef Full Text | Google Scholar
78. Bussmann C, Xia J, Allam JP, Maintz L, Bieber T, Novak N. Early markers for protective mechanisms during rush venom immunotherapy. Allergy. (2010) 65:1558–65. doi: 10.1111/j.1398-9995.2010.02430.x
PubMed Abstract | CrossRef Full Text | Google Scholar
79. Lesourd B, Paupe J, Thiollet M, Moulias R, Sainte-Laudy J, Scheinmann P. Hymenoptera venom immunotherapy. I. Induction of T cell-mediated immunity by honeybee venom immunotherapy: relationships with specific antibody responses. J Allergy Clin Immunol. (1989) 83:563–71. doi: 10.1016/0091-6749(89)90067-5
PubMed Abstract | CrossRef Full Text | Google Scholar
80. Mamessier E, Birnbaum J, Dupuy P, Vervloet D, Magnan A. Ultra-rush venom immunotherapy induces differential T cell activation and regulatory patterns according to the severity of allergy. Clin Exp Allergy. (2006) 36:704–13. doi: 10.1111/j.1365-2222.2006.02487.x
PubMed Abstract | CrossRef Full Text | Google Scholar
81. Schuerwegh AJ, De Clerck LS, Bridts CH, Stevens WJ. Wasp venom immunotherapy induces a shift from IL-4-producing towards interferon-gamma-producing CD4+ and CD8+ T lymphocytes. Clin Exp Allergy. (2001) 31:740–6. doi: 10.1046/j.1365-2222.2001.01066.x
PubMed Abstract | CrossRef Full Text | Google Scholar
82. Tilmant L, Dessaint JP, Tsicopoulos A, Tonnel AB, Capron A. Concomitant augmentation of CD4+ CD45R+ suppressor/inducer subset and diminution of CD4+ CDw29+ helper/inducer subset during rush hyposensitization in hymenoptera venom allergy. Clin Exp Immunol. (1989) 76:13–8.
PubMed Abstract | Google Scholar
83. Shamji MH, Ljorring C, Francis JN, Calderon MA, Larche M, Kimber I, et al. Functional rather than immunoreactive levels of IgG4 correlate closely with clinical response to grass pollen immunotherapy. Allergy. (2012) 67:217–26. doi: 10.1111/j.1398-9995.2011.02745.x
PubMed Abstract | CrossRef Full Text | Google Scholar
84. Shamji MH, Ljorring C, Wurtzen PA. Predictive biomarkers of clinical efficacy of allergen-specific immunotherapy: how to proceed. Immunotherapy. (2013) 5:203–6. doi: 10.2217/imt.13.6
PubMed Abstract | CrossRef Full Text | Google Scholar
85. Bohle B, Kinaciyan T, Gerstmayr M, Radakovics A, Jahn-Schmid B, Ebner C. Sublingual immunotherapy induces IL-10-producing T regulatory cells, allergen-specific T-cell tolerance, and immune deviation. J Allergy Clin Immunol. (2007) 120:707–13. doi: 10.1016/j.jaci.2007.06.013
PubMed Abstract | CrossRef Full Text | Google Scholar
86. Jutel M, Akdis M, Blaser K, Akdis CA. Mechanisms of allergen specific immunotherapy–T-cell tolerance and more. Allergy. (2006) 61:796–807. doi: 10.1111/j.1398-9995.2006.01175.x
PubMed Abstract | CrossRef Full Text | Google Scholar
87. Chen W, Jin W, Hardegen N, Lei KJ, Li L, Marinos N, et al. Conversion of peripheral CD4+CD25− naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J Exp Med. (2003) 198:1875–86. doi: 10.1084/jem.20030152
PubMed Abstract | CrossRef Full Text | Google Scholar
88. Klunker S, Chong MM, Mantel PY, Palomares O, Bassin C, Ziegler M, et al. Transcription factors RUNX1 and RUNX3 in the induction and suppressive function of Foxp3+ inducible regulatory T cells. J Exp Med. (2009) 206:2701–15. doi: 10.1084/jem.20090596
PubMed Abstract | CrossRef Full Text | Google Scholar
89. Meiler F, Zumkehr J, Klunker S, Ruckert B, Akdis CA, Akdis M. In vivo switch to IL-10-secreting T regulatory cells in high dose allergen exposure. J Exp Med. (2008) 205:2887–98. doi: 10.1084/jem.20080193
PubMed Abstract | CrossRef Full Text | Google Scholar
90. Fallarino F, Grohmann U. Using an ancient tool for igniting and propagating immune tolerance: IDO as an inducer and amplifier of regulatory T cell functions. Curr Med Chem. (2011) 18:2215–21. doi: 10.2174/092986711795656027
PubMed Abstract | CrossRef Full Text | Google Scholar
91. Akdis CA, Blesken T, Akdis M, Wuthrich B, Blaser K. Role of interleukin 10 in specific immunotherapy. J Clin Invest. (1998) 102:98–106. doi: 10.1172/JCI2250
PubMed Abstract | CrossRef Full Text | Google Scholar
92. Botturi K, Vervloet D, Magnan A. T cells and allergens relationships: are they that specific? Clin Exp Allergy. (2007) 37:1121–3. doi: 10.1111/j.1365-2222.2007.02773.x
PubMed Abstract | CrossRef Full Text | Google Scholar
93. Radulovic S, Jacobson MR, Durham SR, Nouri-Aria KT. Grass pollen immunotherapy induces Foxp3-expressing CD4+ CD25+ cells in the nasal mucosa. J Allergy Clin Immunol. (2008) 121:1467–72, 1472 e1. doi: 10.1016/j.jaci.2008.03.013
PubMed Abstract | CrossRef Full Text | Google Scholar
94. Pereira-Santos MC, Baptista AP, Melo A, Alves RR, Soares RS, Pedro E, et al. Expansion of circulating Foxp3+)D25bright CD4+ T cells during specific venom immunotherapy. Clin Exp Allergy. (2008) 38:291–7. doi: 10.1111/j.1365-2222.2007.02887.x
PubMed Abstract | CrossRef Full Text | Google Scholar
95. Jutel M, Akdis M, Budak F, Aebischer-Casaulta C, Wrzyszcz M, Blaser K, et al. IL-10 and TGF-beta cooperate in the regulatory T cell response to mucosal allergens in normal immunity and specific immunotherapy. Eur J Immunol. (2003) 33:1205–14. doi: 10.1002/eji.200322919
PubMed Abstract | CrossRef Full Text | Google Scholar
96. Nasser SM, Ying S, Meng Q, Kay AB, Ewan PW. Interleukin-10 levels increase in cutaneous biopsies of patients undergoing wasp venom immunotherapy. Eur J Immunol. (2001) 31:3704–13. doi: 10.1002/1521-4141(200112)31:12<3704::AID-IMMU3704>3.0.CO;2-3
PubMed Abstract | CrossRef Full Text | Google Scholar
97. Varricchi G, Harker J, Borriello F, Marone G, Durham SR, Shamji MH. T follicular helper (Tfh) cells in normal immune responses and in allergic disorders. Allergy. (2016) 71:1086–94. doi: 10.1111/all.12878
PubMed Abstract | CrossRef Full Text | Google Scholar
98. Sage PT, Sharpe AH. T follicular regulatory cells. Immunol Rev. (2016) 271:246–59. doi: 10.1111/imr.12411
PubMed Abstract | CrossRef Full Text | Google Scholar
99. Schulten V, Tripple V, Seumois G, Qian Y, Scheuermann RH, Fu Z, et al. Allergen-specific immunotherapy modulates the balance of circulating Tfh and Tfr cells. J Allergy Clin Immunol. (2018) 141:775–777.e6. doi: 10.1016/j.jaci.2017.04.032
PubMed Abstract | CrossRef Full Text | Google Scholar
100. van de Veen W, Stanic B, Yaman G, Wawrzyniak M, Sollner S, Akdis DG, et al. IgG4 production is confined to human IL-10-producing regulatory B cells that suppress antigen-specific immune responses. J Allergy Clin Immunol. (2013) 131:1204–12. doi: 10.1016/j.jaci.2013.01.014
PubMed Abstract | CrossRef Full Text | Google Scholar
101. Rosser EC, Mauri C. Regulatory B cells: origin, phenotype, and function. Immunity. (2015) 42:607–12. doi: 10.1016/j.immuni.2015.04.005
PubMed Abstract | CrossRef Full Text | Google Scholar
102. Deniz G, van de Veen W, Akdis M. Natural killer cells in patients with allergic diseases. J Allergy Clin Immunol. (2013) 132:527–35. doi: 10.1016/j.jaci.2013.07.030
PubMed Abstract | CrossRef Full Text | Google Scholar
103. Lao-Araya M, Steveling E, Scadding GW, Durham SR, Shamji MH. Seasonal increases in peripheral innate lymphoid type 2 cells are inhibited by subcutaneous grass pollen immunotherapy. J Allergy Clin Immunol. (2014) 134:1193–5.e4. doi: 10.1016/j.jaci.2014.07.029
PubMed Abstract | CrossRef Full Text | Google Scholar
104. Lombardi V, Beuraud C, Neukirch C, Moussu H, Morizur L, Horiot S, et al. Circulating innate lymphoid cells are differentially regulated in allergic and nonallergic subjects. J Allergy Clin Immunol. (2016) 138:305–8. doi: 10.1016/j.jaci.2015.12.1325
PubMed Abstract | CrossRef Full Text | Google Scholar
105. Novak N, Mete N, Bussmann C, Maintz L, Bieber T, Akdis M, et al. Early suppression of basophil activation during allergen-specific immunotherapy by histamine receptor 2. J Allergy Clin Immunol. (2012) 130:1153–8.e2. doi: 10.1016/j.jaci.2012.04.039
PubMed Abstract | CrossRef Full Text | Google Scholar
106. Maintz L, Bussmann C, Bieber T, Novak N. Contribution of histamine metabolism to tachyphylaxis during the buildup phase of rush immunotherapy. J Allergy Clin Immunol. (2009) 123:701–3. doi: 10.1016/j.jaci.2008.11.009
PubMed Abstract | CrossRef Full Text | Google Scholar
107. Cavkaytar O, Akdis CA, Akdis M. Modulation of immune responses by immunotherapy in allergic diseases. Curr Opin Pharmacol. (2014) 17:30–7. doi: 10.1016/j.coph.2014.07.003
PubMed Abstract | CrossRef Full Text | Google Scholar
108. Akdis CA, Jutel M, Akdis M. Regulatory effects of histamine and histamine receptor expression in human allergic immune responses. Chem Immunol Allergy. (2008) 94:67–82. doi: 10.1159/000154858
PubMed Abstract | CrossRef Full Text | Google Scholar
109. Jutel M, Akdis M, Akdis CA. Histamine, histamine receptors and their role in immune pathology. Clin Exp Allergy. (2009) 39:1786–800. doi: 10.1111/j.1365-2222.2009.03374.x
PubMed Abstract | CrossRef Full Text | Google Scholar
110. Mazzoni A, Young HA, Spitzer JH, Visintin A, Segal DM. Histamine regulates cytokine production in maturing dendritic cells, resulting in altered T cell polarization. J Clin Invest. (2001) 108:1865–73. doi: 10.1172/JCI200113930
PubMed Abstract | CrossRef Full Text | Google Scholar
111. Osna N, Elliott K, Khan MM. Regulation of interleukin-10 secretion by histamine in TH2 cells and splenocytes. Int Immunopharmacol. (2001) 1:85–96. doi: 10.1016/S0162-3109(00)00268-X
PubMed Abstract | CrossRef Full Text | Google Scholar
112. Jutel M, Watanabe T, Klunker S, Akdis M, Thomet OA, Malolepszy J, et al. Histamine regulates T-cell and antibody responses by differential expression of H1 and H2 receptors. Nature. (2001) 413:420–5. doi: 10.1038/35096564
PubMed Abstract | CrossRef Full Text | Google Scholar
113. Goldberg A, Confino-Cohen R. Bee venom immunotherapy–how early is it effective? Allergy. (2010) 65:391–5. doi: 10.1111/j.1398-9995.2009.02198.x
PubMed Abstract | CrossRef Full Text | Google Scholar
114. Frick M, Fischer J, Helbling A, Rueff F, Wieczorek D, Ollert M, et al. Predominant Api m 10 sensitization as risk factor for treatment failure in honey bee venom immunotherapy. J Allergy Clin Immunol. (2016) 138:1663–71.e9. doi: 10.1016/j.jaci.2016.04.024
PubMed Abstract | CrossRef Full Text | Google Scholar
115. Blank S, Seismann H, Michel Y, McIntyre M, Cifuentes L, Braren I, et al. Api m 10, a genuine A. mellifera venom allergen, is clinically relevant but underrepresented in therapeutic extracts. Allergy. (2011) 66:1322–9. doi: 10.1111/j.1398-9995.2011.02667.x
PubMed Abstract | CrossRef Full Text | Google Scholar
116. Stretz E, Oppel EM, Rawer HC, Chatelain R, Mastnik S, Przybilla B, et al. Overcoming severe adverse reactions to venom immunotherapy using anti-IgE antibodies in combination with a high maintenance dose. Clin Exp Allergy. (2017) 47:1631–9. doi: 10.1111/cea.12997
PubMed Abstract | CrossRef Full Text | Google Scholar
117. Bousquet J, Menardo JL, Velasquez G, Michel FB. Systemic reactions during maintenance immunotherapy with honey bee venom. Ann Allergy. (1988) 61:63–8.
PubMed Abstract | Google Scholar
118. Rueff F, Wenderoth A, Przybilla B. Patients still reacting to a sting challenge while receiving conventional Hymenoptera venom immunotherapy are protected by increased venom doses. J Allergy Clin Immunol. (2001) 108:1027–32. doi: 10.1067/mai.2001.119154
PubMed Abstract | CrossRef Full Text | Google Scholar
119. Golden DB, Johnson K, Addison BI, Valentine MD, Kagey-Sobotka A, Lichtenstein LM. Clinical and immunologic observations in patients who stop venom immunotherapy. J Allergy Clin Immunol. (1986) 77:435–42. doi: 10.1016/0091-6749(86)90177-6
PubMed Abstract | CrossRef Full Text | Google Scholar
120. Muller UR, Ring J. When can immunotherapy for insect sting allergy be stopped? J Allergy Clin Immunol Pract. (2015) 3:324–8. doi: 10.1016/j.jaip.2014.11.018
PubMed Abstract | CrossRef Full Text | Google Scholar
121. Golden DB, Kwiterovich KA, Kagey-Sobotka A, Valentine MD, Lichtenstein LM. Discontinuing venom immunotherapy: outcome after five years. J Allergy Clin Immunol. (1996) 97:579–87. doi: 10.1016/S0091-6749(96)70302-0
PubMed Abstract | CrossRef Full Text | Google Scholar
122. Dhami S, Zaman H, Varga EM, Sturm GJ, Muraro A, Akdis CA, et al. Allergen immunotherapy for insect venom allergy: a systematic review and meta-analysis. Allergy. (2017) 72:342–65. doi: 10.1111/all.13077
PubMed Abstract | CrossRef Full Text | Google Scholar
123. Korosec P, Ziberna K, Silar M, Dezman M, Celesnik Smodis N, Rijavec M, et al. Immunological and clinical factors associated with adverse systemic reactions during the build-up phase of honeybee venom immunotherapy. Clin Exp Allergy. (2015) 45:1579–89. doi: 10.1111/cea.12582
PubMed Abstract | CrossRef Full Text | Google Scholar
124. Golden DB. Long-term outcome after venom immunotherapy. Curr Opin Allergy Clin Immunol. (2010) 10:337–41. doi: 10.1097/ACI.0b013e32833bc0ba
PubMed Abstract | CrossRef Full Text | Google Scholar
125. Muller U, Berchtold E, Helbling A. Honeybee venom allergy: results of a sting challenge 1 year after stopping successful venom immunotherapy in 86 patients. J Allergy Clin Immunol. (1991) 87:702–9. doi: 10.1016/0091-6749(91)90392-2
PubMed Abstract | CrossRef Full Text | Google Scholar
126. Rueff F, Przybilla B, Bilo MB, Muller U, Scheipl F, Seitz MJ, et al. Clinical effectiveness of hymenoptera venom immunotherapy: a prospective observational multicenter study of the European academy of allergology and clinical immunology interest group on insect venom hypersensitivity. PLoS ONE. (2013) 8:e63233. doi: 10.1371/journal.pone.0063233
PubMed Abstract | CrossRef Full Text | Google Scholar
127. Slade CA, Douglass JA. Changing practice: no need to stop ACE inhibition for venom immunotherapy. Clin Exp Allergy. (2014) 44:617–9. doi: 10.1111/cea.12295
PubMed Abstract | CrossRef Full Text | Google Scholar
128. Golden DB, Kwiterovich KA, Kagey-Sobotka A, Lichtenstein LM. Discontinuing venom immunotherapy: extended observations. J Allergy Clin Immunol. (1998) 101:298–305. doi: 10.1016/S0091-6749(98)70239-8
PubMed Abstract | CrossRef Full Text | Google Scholar
129. Mobs C, Muller J, Rudzio A, Pickert J, Blank S, Jakob T, et al. Decline of Ves v 5-specific blocking capacity in wasp venom-allergic patients after stopping allergen immunotherapy. Allergy. (2015) 70:715–9. doi: 10.1111/all.12606
PubMed Abstract | CrossRef Full Text | Google Scholar
Zdroj: https://www.frontiersin.org/articles/10.3389/fimmu.2019.01959/full